La neuroplasticité – l'extraordinaire capacité du cerveau à se réorganiser, à modifier ses connexions et à créer de nouveaux circuits neuronaux – constitue le mécanisme fondamental sous-jacent à tout apprentissage, à toute récupération et à toute optimisation cognitive. Chez Neurofeedback Luxembourg, nous comprenons que maximiser la neuroplasticité nécessite plus qu'un entraînement ; elle exige la création d'un environnement cellulaire optimal. Photobiomodulation transcrânienne (tPBM) par Vielight la technologie offre une approche révolutionnaire pour activer et amplifier les mécanismes neuroplastiques à leur source.
Le tPBM utilise des sources lumineuses non ionisantes du spectre électromagnétique pour interagir avec le corps humain au niveau cellulaire.
Introduction à la neuroplasticité et au tPBM
La neuroplasticité est la remarquable capacité du cerveau à se réorganiser en formant de nouvelles connexions neuronales tout au long de la vie. Ce processus adaptatif permet au cerveau de réagir aux nouvelles expériences, de se rétablir des blessures et d'améliorer l'apprentissage et la mémoire. La photobiomodulation transcrânienne (tPBM) est une technique innovante et non invasive qui exploite la luminothérapie de faible intensité, notamment la thérapie laser de faible intensité et les diodes électroluminescentes (DEL), pour stimuler la capacité naturelle du cerveau à s'adapter. En délivrant une lumière proche infrarouge au cuir chevelu, la tPBM cible la cytochrome c oxydase, une enzyme clé de la chaîne respiratoire mitochondriale. Cette interaction stimule le potentiel de la membrane mitochondriale, augmente la production d'ATP et génère des niveaux contrôlés d'espèces réactives de l'oxygène (ERO), toutes essentielles au métabolisme et à la signalisation cellulaires. Des études cliniques ont montré que la tPBM peut améliorer significativement les fonctions cognitives, en particulier chez les personnes souffrant de traumatisme crânien, de trouble dépressif majeur et d'autres affections neurologiques. La capacité du tPBM à moduler l’activité mitochondriale et la production de ROS sous-tend son potentiel à stimuler la neuroplasticité et à soutenir la santé du cerveau.
Comprendre la neuroplasticité : les mécanismes fondamentaux
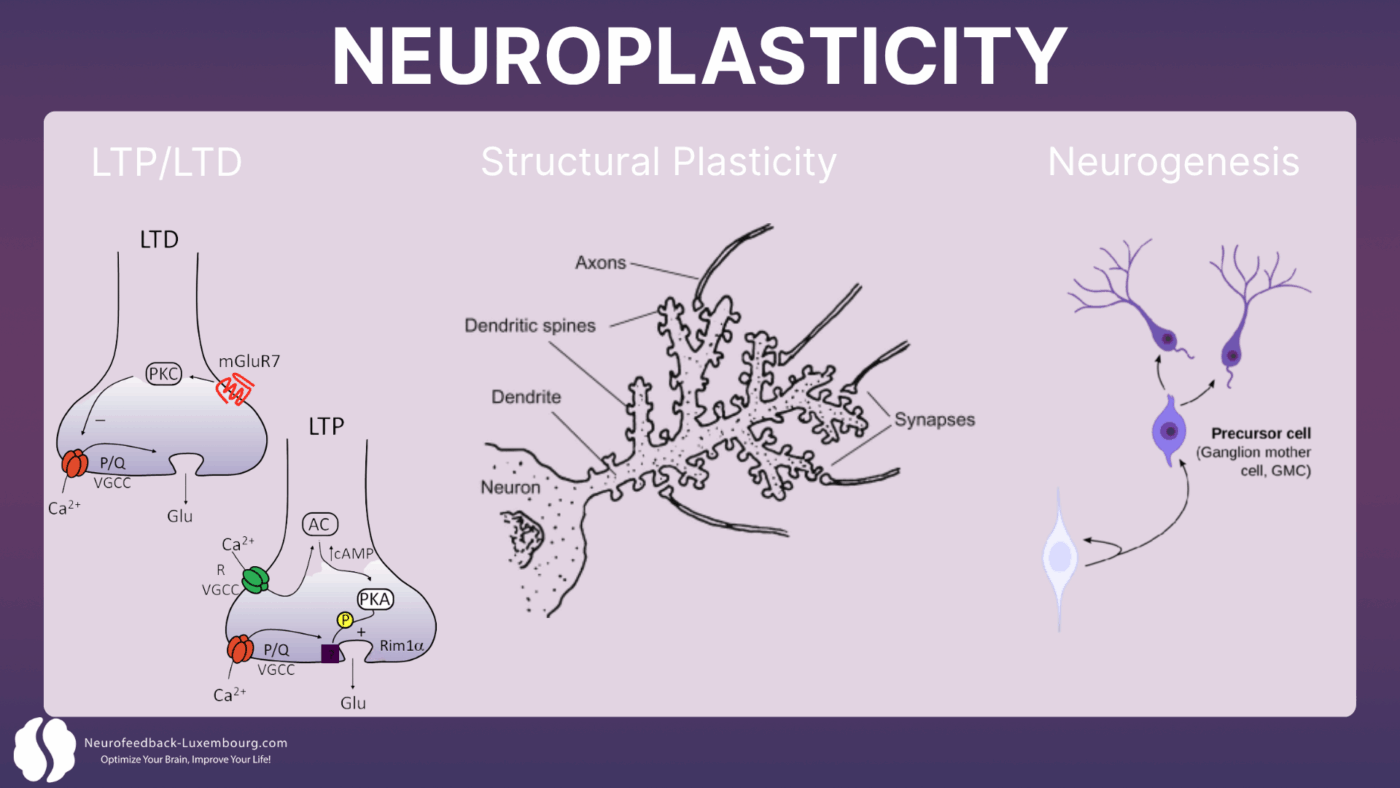
Plasticité synaptique : le fondement de l'adaptation
La neuroplasticité opère principalement à travers quatre mécanismes interconnectés:
1. Potentialisation à long terme (LTP):Le renforcement durable des connexions synaptiques
2. Dépression à long terme (LTD):L'affaiblissement sélectif des connexions inutilisées
3. Plasticité structurelle:La formation de nouvelles épines dendritiques et de synapses
4. Neurogenèse:La création de nouveaux neurones dans des régions spécifiques du cerveau
Ces processus nécessitent synchronisation précise d'activité électrique, d'une disponibilité énergétique substantielle et d'un environnement biochimique optimal. C'est précisément là qu'intervient le tPBM en tant que catalyseur neuroplastique.
Énergie lumineuse et fonction cérébrale
L'énergie lumineuse, notamment dans le spectre proche infrarouge, joue un rôle essentiel dans l'optimisation des fonctions cérébrales. Appliquée par luminothérapie de faible intensité, la lumière proche infrarouge pénètre les tissus biologiques et interagit avec les composants cellulaires pour déclencher une cascade d'effets bénéfiques (1)(2). Des recherches ont démontré que le tPBM peut augmenter le flux sanguin cérébral, favorisant ainsi l'apport d'oxygène et de nutriments aux régions cérébrales actives (3)(4). Cette perfusion améliorée, combinée à la régulation positive de facteurs de croissance tels que le facteur de croissance de l'endothélium vasculaire (VEGF), favorise la réparation et la régénération tissulaires (5). De plus, il a été démontré que le tPBM stimule la neurogenèse et la plasticité synaptique, deux éléments essentiels à l'apprentissage et à la mémoire (6). La réduction du stress oxydatif et de l'inflammation contribue également à un environnement neuronal sain. Pour des résultats optimaux, les protocoles tPBM utilisent généralement des densités de puissance comprises entre 10 et 100 mW/cm² et des doses d'énergie comprises entre 1 et 10 J/cm², garantissant un apport lumineux efficace aux tissus cibles tout en maximisant la sécurité et l'efficacité. Ces paramètres sont essentiels pour exploiter tout le potentiel de la luminothérapie pour améliorer la fonction cérébrale et favoriser la réparation des tissus.
Apport de lumière au cerveau
L'administration efficace de lumière au cerveau est essentielle à la réussite de la thérapie tPBM. Plusieurs approches ont été développées pour garantir que la lumière proche infrarouge atteigne les tissus cibles. La tPBM transcrânienne consiste à appliquer la lumière infrarouge directement sur le cuir chevelu, permettant une pénétration profonde de la lumière dans les régions corticales et sous-corticales. La tPBM intranasale diffuse la lumière par les fosses nasales, permettant ainsi l'accès aux structures cérébrales via le riche réseau vasculaire de cette zone. Les méthodes extracrâniennes, comme l'application de lumière au niveau du cou ou du torse, peuvent stimuler des effets systémiques bénéfiques indirects pour la santé cérébrale. Le choix de la méthode d'administration dépend de l'affection neurologique spécifique et des objectifs thérapeutiques. Il est important de noter que la lumière proche infrarouge est privilégiée par rapport à la lumière visible en raison de sa meilleure pénétration tissulaire, lui permettant d'atteindre des structures cérébrales plus profondes, inaccessibles aux longueurs d'onde plus courtes (2). Comprendre les principes de pénétration de la lumière et les propriétés uniques de la lumière proche infrarouge est essentiel pour optimiser les résultats de la tPBM.
Comment le tPBM active directement la neuroplasticité
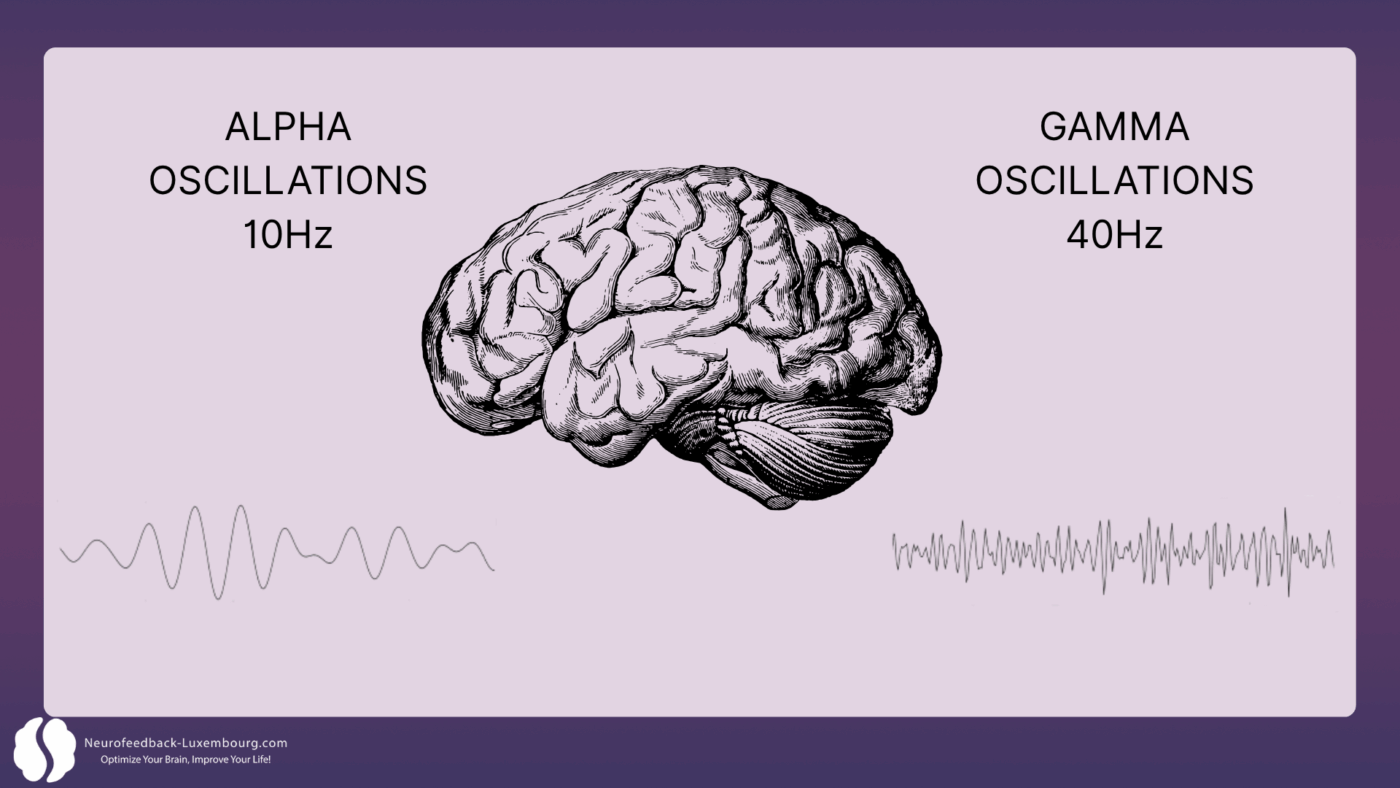
Modulation de l'oscillation neuronale : le timing de la plasticité
La neuroplasticité dépend fondamentalement de la timing précis de l'activité neuronale. Des études démontrent que le tPBM influence directement les oscillations cérébrales qui orchestrent les changements plastiques :
Oscillations gamma (40 Hz) et LTP:
- Il a été démontré que la stimulation à 40 Hz améliore la mémoire et l'apprentissage, tout en réduisant les plaques amyloïdes (1). Cette modulation est supposée augmenter la microglie non inflammatoire, responsable de l'élimination des plaques amyloïdes. L'administration d'énergie lumineuse proche infrarouge pulsée à 40 Hz augmente significativement la puissance des ondes gamma tout en réduisant les fréquences plus lentes, favorisant ainsi un couplage thêta-gamma renforcé, corrélé à une amélioration de l'apprentissage spatial et des performances cognitives.
- Résultat : renforcement accéléré des connexions synaptiques lors de l’apprentissage.
Oscillations alpha (10 Hz) et consolidation:
- Les rythmes alpha (10 Hz) favorisent la pleine conscience, l'apprentissage et la relaxation. Le mode alpha 10 Hz a montré des résultats supérieurs à ceux du mode 40 Hz lors d'études cliniques, avec des résultats cognitifs nettement supérieurs après 12 semaines de traitement. Le PBM peut influencer le couplage hippocampique-cortical, essentiel à l'apprentissage et à la mémoire (1).
- Résultats : Amélioré facilitation de l'apprentissage et la stabilisation des états détendus et concentrés.
Vielight la recherche démontre que le tPBM entraîne directement ces fréquences, créant des fenêtres temporelles optimales pour induire des changements plastiques.
Activation des voies de signalisation plastique
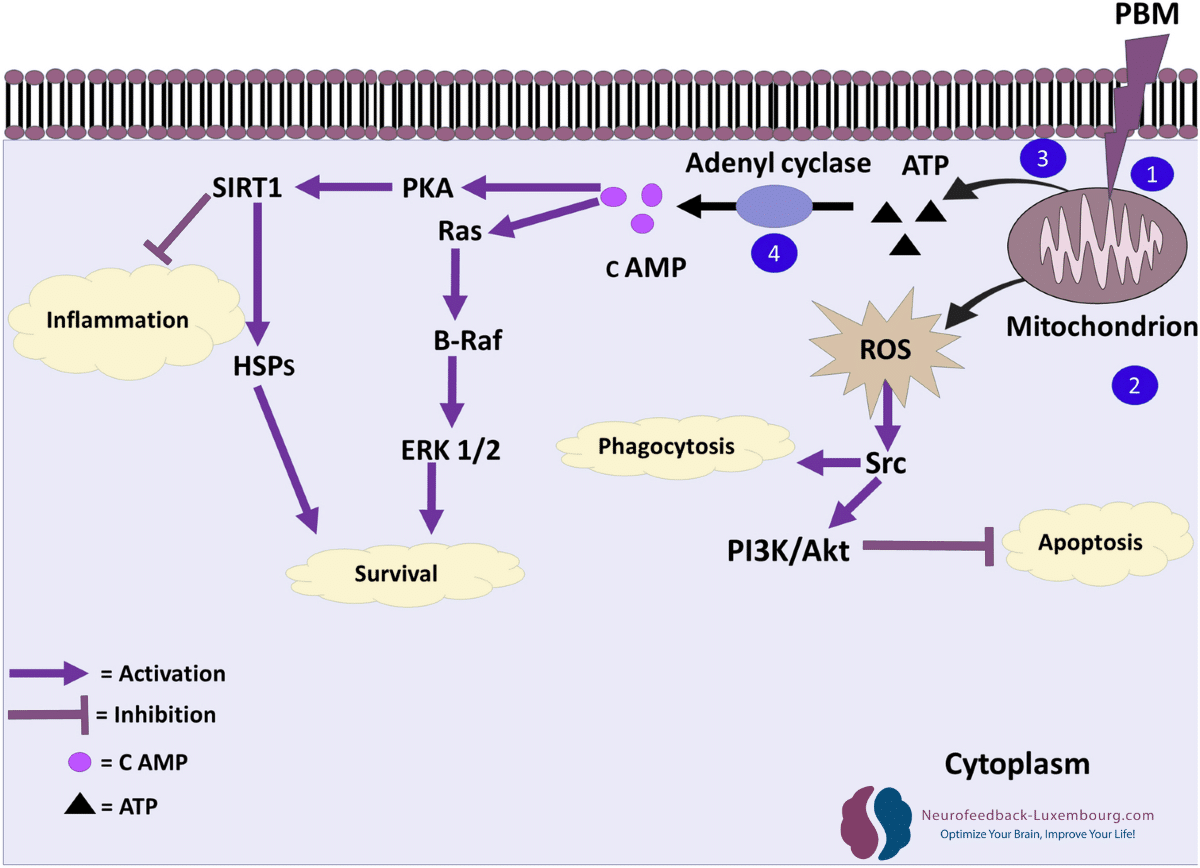
Le tPBM déclenche spécifiquement les cascades moléculaires qui régissent la neuroplasticité :
Voie Akt/GSK3β/β-caténine:
- Activation directe par stimulation mitochondriale
- Promotion de survie neuronale et la croissance dendritique
- Facilitation de plasticité structurelle
Signalisation CREB (liaison de l'élément de réponse à l'AMPc):
- Phosphorylation CREB améliorée grâce à l'augmentation de l'ATP
- Régulation positive de gènes de plasticité (Arc, c-Fos, Zif268)(7) (8)
- Synthèse accélérée des protéines nécessaires aux changements synaptiques
Le tPBM peut également influencer les facteurs de transcription tels que AP-1 et NF-κB, qui régulent l'expression des gènes en réponse aux signaux cellulaires.
Cascade mTOR (cible mécanistique de la rapamycine):
- Activation par un métabolisme énergétique amélioré
- Réglementation de synthèse protéique locale aux synapses
- Contrôle de la croissance dendritique et de la formation des épines (9)
Au-delà de ces mécanismes moléculaires, le tPBM peut activer les canaux ioniques sensibles à la lumière, y compris les canaux de potentiel récepteur transitoire, qui contribuent à ses effets sur la signalisation neuronale.
BDNF : le régulateur principal de la neuroplasticité
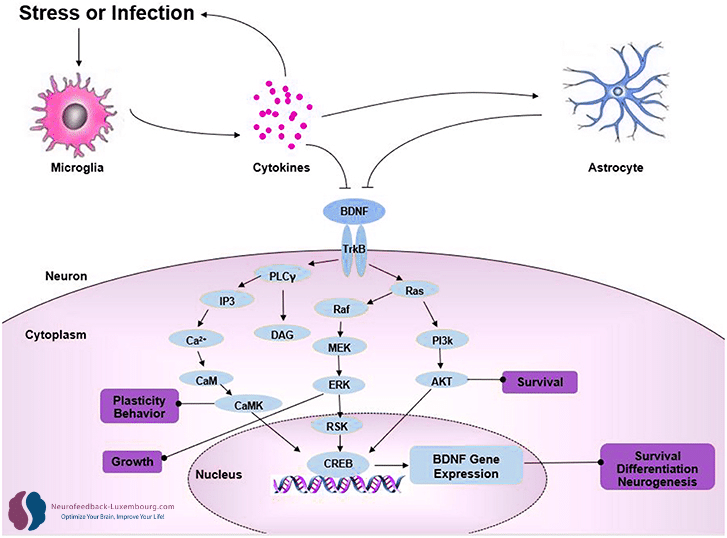
Régulation positive massive par tPBM
Le facteur neurotrophique dérivé du cerveau (BDNF) orchestre pratiquement tous les aspects de la neuroplasticité. Le tPBM produit un augmentation spectaculaire dans les niveaux de BDNF jusqu'à (10) :
Mécanismes transcriptionnels:
- Activation du promoteur BDNF via CREB(11) phosphorylé
- Expression améliorée de l'ARNm du BDNF dans l'hippocampe et le cortex
- Augmentation de la synthèse de pro-BDNF et de BDNF mature
Effets fonctionnels directs:
- Plasticité de la colonne vertébrale: Promotion de la formation et de la maturation des épines dendritiques
- Force synaptique:Facilitation de l'insertion du récepteur AMPA
- Plasticité structurelle:Stimulation de la croissance axonale et dendritique
BDNF et plasticité de la période critique
La recherche révèle que le tPBM peut rouvrir les fenêtres critiques de la plasticité chez l'adulte (12) :
- Réactivation des mécanismes de plasticité juvénile
- Réduction des réseaux périneuronaux limitant la plasticité
- Restauration de plasticité de dominance oculaire dans les modèles animaux
Cette capacité suggère que le tPBM peut surmonter les limitations liées à l'âge de la neuroplasticité.
Neurogenèse adulte : création de nouveaux neurones
Stimulation directe des niches neurogènes
Contrairement aux approches qui modulent simplement l’activité neuronale existante, le tPBM stimule la création de nouveaux neurones dans les modèles animaux :
Zone sous-ventriculaire (ZSV) (13):
- Prolifération accrue des cellules souches neurales
- Amélioration de la migration vers les régions cibles
- Différenciation accrue en neurones fonctionnels
Gyrus denté de l'hippocampe:
- Activation des cellules souches de type 1 (de type glie radiale)
- Promotion de la maturation des neurones granuleux
- Intégration fonctionnelle dans les circuits existants
Mécanismes moléculaires:
- Activation du TGF-β1 latent favorisant la neurogenèse (14)(15)
- Signalisation Wnt améliorée favorisant la prolifération des cellules souches
- Inhibition inflammatoire réduite de la neurogenèse
Impact fonctionnel de la neurogenèse
Les nouveaux neurones générés par tPBM contribuent spécifiquement à :
- Séparation des motifs:Discrimination entre souvenirs similaires (16)
- Flexibilité cognitive:Adaptation à de nouveaux environnements (17)
- Régulation de l'humeur: Modulation des circuits émotionnels
- Capacité d'apprentissage:Acquisition améliorée de nouvelles compétences
Synaptogenèse : construire de nouveaux circuits
Formation accélérée de synapses
tPBM influence directement synaptogenèse—la formation de nouvelles connexions synaptiques :
Germination axonale:
- Stimulation de l'avancement des cônes de croissance
- Expression améliorée des protéines de guidage axonal
- Facilitation de la formation de nouveaux boutons synaptiques
Formation d'épines dendritiques:
- Augmentation de la densité de la colonne vertébrale dans les études animales (18)
- Maturation améliorée des épines immatures
- Stabilisation de nouvelles connexions grâce à une activité coordonnée
Assemblage du complexe synaptique:
- Régulation positive des protéines d'échafaudage (PSD-95, SHANK)
- Regroupement amélioré des récepteurs de neurotransmetteurs
- Amélioration du trafic vésiculaire et de la libération des neurotransmetteurs
Spécificité dépendante du circuit
Il est remarquable que le tPBM influence la synaptogenèse dans un spécifique au circuit manière:
- Connectivité améliorée dans les réseaux actif pendant le traitement
- Renforcement sélectif de fonctionnellement pertinent relations
- Amélioration de l'élagage des connexions inutilisées via LTD
Cette spécificité explique pourquoi tPBM potentialise effets de neurofeedback sans créer de changements aléatoires.
Plasticité homéostatique : maintenir l'équilibre
Régulation de l'excitabilité neuronale
Influences du tPBM plasticité homéostatique mécanismes qui maintiennent la stabilité du réseau :
Mise à l'échelle synaptique:
- Ajustement global de la force synaptique
- Prévention de l'hyperexcitabilité ou de l'hypoexcitabilité
- Maintien de plages dynamiques optimales
Plasticité intrinsèque:
- Modulation des canaux ioniques voltage-dépendants
- Ajustement du seuil d'excitation neuronale
- Optimisation de l'intégration dendritique
Ce autorégulation garantit que les changements plastiques induits restent dans des limites physiologiques saines.
Fondation pour l'énergie cellulaire : rôle de la cytochrome c oxydase
Production d'ATP améliorée : alimenter l'adaptation neuronale
Les changements neuroplastiques sont des processus fondamentalement énergivores. La formation de nouvelles synapses, la ramification dendritique et le remodelage de la myéline nécessitent d'importantes ressources énergétiques cellulaires. La tPBM répond directement à ce besoin grâce à stimulation de la cytochrome c oxydase—l'enzyme photosensible du complexe IV de la chaîne de transport d'électrons mitochondriaux.
Lorsque la lumière proche infrarouge (810 nm) pénètre dans le tissu cérébral, elle cible spécifiquement cette enzyme mitochondriale, déclenchant une cascade d'effets bioénergétiques :
- Augmentation immédiate de l'ATP:Des études démontrent une augmentation significative de la production d'adénosine triphosphate dans les minutes suivant l'application de tPBM
- Phosphorylation oxydative améliorée:L'amélioration de l'efficacité de la chaîne de transport des électrons favorise une production d'énergie durable
- Optimisation du potentiel de la membrane mitochondriale: Restauration de gradients électriques optimaux à travers les membranes mitochondriales
Ce amélioration métabolique crée un environnement cellulaire idéal où les neurones possèdent les ressources énergétiques nécessaires pour soutenir une adaptation et un apprentissage rapides.
Amélioration de la microcirculation : le fondement vasculaire
Au-delà de la production directe d'énergie, le tPBM stimule la synthèse d'oxyde nitrique (NO), produisant des améliorations mesurables dans :
- Débit sanguin cérébral:La perfusion régionale augmente après les séances de tPBM
- apport d'oxygène:Saturation de l'hémoglobine et oxygénation des tissus améliorées dans les régions traitées
- Transport des nutriments: Amélioration de l'apport de glucose, d'acides aminés et de facteurs neurotrophiques aux réseaux neuronaux actifs
Les recherches démontrent une augmentation significative de la perfusion cérébrale dans les lobes frontaux, temporaux et occipitaux, ainsi que dans l'hippocampe, après un traitement par tPBM. Ces améliorations vasculaires créent une microenvironnement riche en nutriments qui soutient une activité neuroplastique soutenue.
Neuroprotection anti-inflammatoire
Production contrôlée d'espèces réactives de l'oxygène (ROS)
Alors que le stress oxydatif excessif endommage le tissu nerveux, le tPBM produit espèces réactives de l'oxygène (ROS) contrôlées et douces qui fonctionnent comme des molécules de signalisation bénéfiques. Cette réponse hormétique déclenche :
- Régulation positive des enzymes antioxydantes:Activité améliorée de la catalase, de la superoxyde dismutase et de la glutathion peroxydase
- Production de cytokines anti-inflammatoires:Augmentation de l'IL-10 et diminution des marqueurs pro-inflammatoires (TNF-α, IL-1β)
- Modulation du phénotype microglial:Passage de l'activation microgliale M1 (inflammatoire) à M2 (réparatrice)
Ce environnement neuroprotecteur prévient les dommages cellulaires qui entravent généralement les processus neuroplastiques en cas de stress ou après une lésion neuronale. Intégration clinique : maximiser la neuroplasticité
Synergique Protocoles tPBM-Neurofeedback
L'intégration optimale exploite les fenêtres temporelles de la neuroplasticité :
Phase de préparation (tPBM 20 min):
- Vielight Neuro Gamma (40 Hz) : Activation des réseaux attentionnels
- Amorçage des mécanismes LTP pour l'apprentissage ultérieur
- Régulation rapide du BDNF et des facteurs plastiques
Phase d'entraînement (Neurofeedback):
- Application de protocoles guidés par QEEG pendant une période optimale fenêtre en plastique
- Utilisation de la synchronisation neuronale améliorée
- Exploitation de l'excitabilité contrôlée accrue
Phase de consolidation (tPBM 20 min):
- Vielight Neuro Alpha (10 Hz) : Facilitation de la consolidation de la mémoire
- Stabilisation des modèles nouvellement appris
- Synthèse protéique améliorée favorisant les changements structurels
Biomarqueurs de neuroplasticité
Nous surveillons l'efficacité au moyen de mesures spécifiques indicateurs neuroplastiques:
Marqueurs QEEG:
- Cohérence accrue dans les bandes de fréquences entraînées
- Couplage interfréquence amélioré (thêta-gamma)
- Amélioration des mesures d'efficacité du réseau
Évaluations comportementales:
- Courbes d'acquisition accélérées pendant la formation
- Amélioration de la rétention entre les sessions
- Généralisation améliorée aux tâches non entraînées
Conclusion : La révolution neuroplastique
La photobiomodulation transcrânienne représente une changement de paradigme dans notre approche de la neuroplasticité. Plutôt que de simplement stimulant le cerveau à changer, tPBM active les mécanismes fondamentaux qui rendent tout changement neuroplastique possible.
Cette technologie nous permet de :
- Rouvrir les fenêtres critiques de la plasticité à tout âge
- Accélérer les processus de LTP, de neurogenèse et de synaptogenèse
- Optimiser le timing des interventions neuroplastiques
- Maximiser la rétention et la généralisation des changements
Pour les praticiens du neurofeedback, le tPBM offre l’opportunité unique de travailler avec la neuroplasticité plutôt que de s'opposer à ses limites naturelles. Résultat : des changements plus rapides, plus durables et plus profonds que jamais.
Découvrez dès aujourd'hui votre potentiel d'optimisation cérébrale. Visitez notre site web pour en savoir plus. neurofeedback et notre approche intégrée, et planifiez votre téléconsultation préliminaireEnsemble, explorons comment la photobiomodulation transcrânienne peut transformer la santé de votre cerveau.
L'avenir de l'optimisation cérébrale réside dans la compréhension et l'activation directe des mécanismes neuroplastiques. Chez Neurofeedback Luxembourg, nous combinons notre expertise en EEGQ avec la technologie Vielight pour créer des programmes d'entraînement cérébral performants. informé neurologiquement et optimisé neuroplastiquement.
Tous les protocoles sont conçus pour optimiser votre bien-être et ne constituent pas un traitement médical. Les résultats peuvent varier d'une personne à l'autre. Une consultation est recommandée pour une évaluation personnalisée.
Références
(2) Naeser, MA, Martin, PI, Ho, MD, Krengel, MH, Bogdanova, Y., Knight, JA, Hamblin, MR, Fedoruk, AE, Poole, LG, Cheng, C., & Koo, B. (sd). Traitement par photobiomodulation transcrânienne : améliorations significatives chez quatre anciens joueurs de football atteints d'une possible encéphalopathie traumatique chronique. Journal des rapports sur la maladie d'Alzheimer, 7(1), 77‑105. https://doi.org/10.3233/ADR-220022
(1) Lim, L. (2024). Modification de la physiopathologie de la maladie d'Alzheimer par photobiomodulation : modèle, preuves et avenir avec une intervention guidée par EEG. Frontières en neurologie, 15. https://doi.org/10.3389/fneur.2024.1407785
(3) Chao, LL (2019). Effets des traitements de photobiomodulation à domicile sur la fonction cognitive et comportementale, la perfusion cérébrale et la connectivité fonctionnelle à l'état au repos chez les patients atteints de démence: un essai pilote. Photobiomodulation, photomédicine et chirurgie laser, 37(3), 133‑141. https://doi.org/10.1089/photob.2018.4555
(4) Chao, LL, Barlow, C., Karimpoor, M., et Lim, L. (2020). Modifications de la fonction et de la structure cérébrales après un traitement auto-administré de photobiomodulation à domicile dans un cas de commotion cérébrale. Frontières en neurologie, 11. https://doi.org/10.3389/fneur.2020.00952
(5) Di Gregorio, E., Staelens, M., Hosseinkhah, N., Karimpoor, M., Liburd, J., Lim, L., Shankar, K., & Tuszyński, JA (2024). La spectroscopie Raman révèle une transition de l'hélice α au feuillet β induite par photobiomodulation dans les tubulines : implications potentielles pour la maladie d'Alzheimer et d'autres maladies neurodégénératives. Nanomatériaux, 14(13), article 13. https://doi.org/10.3390/nano14131093
(6) Français Lim, L., Hosseinkhah, N., Van Buskirk, M., Berk, A., Loheswaran, G., Abbaspour, Z., Karimpoor, M., Smith, A., Ho, KF, Pushparaj, A., Zahavi, M., White, A., Rubine, J., Zidel, B., Henderson, C., Clayton, RG, Tingley, DR, Miller, DJ, Karimpoor, M., & Hamblin, MR (2024). Traitement par photobiomodulation avec un dispositif à domicile pour la COVID-19 : un essai contrôlé randomisé pour l'efficacité et la sécurité. Photobiomodulation, photomédicine et chirurgie laser, 42(6), 393‑403. https://doi.org/10.1089/pho.2023.0179
(7) Knapska, E., & Kaczmarek, L. (2004). Un gène pour la plasticité neuronale dans le cerveau des mammifères : Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK ? Progrès en neurobiologie, 74(4), 183‑211. https://doi.org/10.1016/j.pneurobio.2004.05.007
(8) Penke, Z., Morice, E., Veyrac, A., Gros, A., Chagneau, C., LeBlanc, P., Samson, N., Baumgärtel, K., Mansuy, IM, Davis, S., & Laroche, S. (2014). Le gain de fonction de Zif268/Egr1 facilite la plasticité synaptique hippocampique et la mémoire de reconnaissance spatiale à long terme. Philosophical Transactions of the Royal Society B : Sciences biologiques, 369(1633), 20130159. https://doi.org/10.1098/rstb.2013.0159
(9) Lee, DY (2015). Rôles de la signalisation mTOR dans le développement cérébral. Neurobiologie expérimentale, 24(3), 177‑185. https://doi.org/10.5607/en.2015.24.3.177
(11) Heo, J.-C., Park, J.-A., Kim, D.-K., & Lee, J.-H. (2019). La thérapie par photobiomodulation (660 nm) réduit le stress oxydatif et induit l'expression du BDNF dans l'hippocampe. Rapports scientifiques, 9(1), 10114. https://doi.org/10.1038/s41598-019-46490-4
(10) de Oliveira, BH, Lins, EF, Kunde, NF, Salgado, ASI, Martins, LM, Bobinski, F., Vieira, WF, Cassano, P., Quialheiro, A., & Martins, DF (2024). La photobiomodulation transcrânienne augmente la cognition et les taux sériques de BDNF chez les adultes de plus de 50 ans : un essai randomisé, en double aveugle, contrôlé par placebo. Journal de photochimie et de photobiologie B : Biologie, 260, 113041. https://doi.org/10.1016/j.jphotobiol.2024.113041
(12) Hensch, TK, et Bilimoria, PM (2012). Réouverture des fenêtres : manipulation des périodes critiques pour le développement cérébral. Cerebrum : le forum Dana sur les sciences du cerveau, 2012, 11.
(13) Chang, S.-Y., et Lee, MY (2023). Photobiomodulation de la neurogenèse par l'amélioration de la différenciation des cellules souches et des progéniteurs neuronaux dans les systèmes nerveux central et périphérique. Revue internationale des sciences moléculaires, 24(20), 15427. https://doi.org/10.3390/ijms242015427
(14) Wu, X., Shen, Q., Zhang, Z., Zhang, D., Gu, Y., & Xing, D. (2021). La photoactivation de la voie de signalisation TGFβ/SMAD améliore la neurogenèse hippocampique adulte dans le modèle de la maladie d'Alzheimer. Recherche et thérapie sur les cellules souches, 12(1), 345. https://doi.org/10.1186/s13287-021-02399-2
(15) Kandasamy, M., Lehner, B., Kraus, S., Sander, PR, Marschallinger, J., Rivera, FJ, Trümbach, D., Ueberham, U., Reitsamer, HA, Strauss, O., Bogdahn, U., Couillard-Despres, S., & Aigner, L. (2014). La signalisation TGF-bêta dans la niche neurogénique adulte favorise la quiescence des cellules souches ainsi que la génération de nouveaux neurones. Journal de médecine cellulaire et moléculaire, 18(7), 1444‑1459. https://doi.org/10.1111/jcmm.12298
(16) Johnston, ST, Shtrahman, M., Parylak, S., Gonçalves, JT, & Gage, FH (2016). Paradoxe de la séparation des motifs et neurogenèse adulte : un double rôle pour les nouveaux neurones équilibrant la résolution de la mémoire et la robustesse. Neurobiologie de l'apprentissage et de la mémoire, 129, 60‑68. https://doi.org/10.1016/j.nlm.2015.10.013
(17) Anacker, C. et Hen, R. (2017). Neurogenèse hippocampique adulte et flexibilité cognitive : lien entre mémoire et humeur. Nature Reviews Neuroscience, 18(6), 335‑346. https://doi.org/10.1038/nrn.2017.45
(18) Meng, C., He, Z., & Xing, D. (2013). La thérapie laser de faible intensité sauve l'atrophie des dendrites en régulant positivement l'expression du BDNF : implications pour la maladie d'Alzheimer. The Journal of Neuroscience : Le journal officiel de la Society for Neuroscience, 33(33), 13505‑13517. https://doi.org/10.1523/JNEUROSCI.0918-13.2013
