Neuroplastizität – die außergewöhnliche Fähigkeit des Gehirns, sich neu zu organisieren, seine Verbindungen zu verändern und neue neuronale Schaltkreise zu bilden – ist der grundlegende Mechanismus, der allen Lernprozessen, der Genesung und der kognitiven Optimierung zugrunde liegt. Wir bei Neurofeedback Luxembourg verstehen das Maximierung der Neuroplastizität Es erfordert mehr als nur Training; es verlangt die Schaffung eines optimalen zellulären Milieus. Transkranielle Photobiomodulation (tPBM) durch Vielight Technologie bietet einen revolutionären Ansatz für neuroplastische Mechanismen aktivieren und verstärken an ihrem Ursprung.
tPBM nutzt nichtionisierende Lichtquellen aus dem elektromagnetischen Spektrum, um auf zellulärer Ebene mit dem menschlichen Körper zu interagieren.
Einführung in die Neuroplastizität und tPBM
Neuroplastizität ist die bemerkenswerte Fähigkeit des Gehirns, sich durch die Bildung neuer neuronaler Verbindungen lebenslang selbst zu reorganisieren. Dieser adaptive Prozess ermöglicht es dem Gehirn, auf neue Erfahrungen zu reagieren, sich von Verletzungen zu erholen und Lernen und Gedächtnis zu verbessern. Die transkranielle Photobiomodulation (tPBM) ist eine innovative, nicht-invasive Technik, die sich die Therapie mit niedrigdosiertem Licht – einschließlich Low-Level-Lasertherapie und Leuchtdioden – zunutze macht, um die natürliche Veränderungsfähigkeit des Gehirns zu stimulieren. Durch die Applikation von Nahinfrarotlicht auf die Kopfhaut zielt tPBM auf die Cytochrom-c-Oxidase ab, ein Schlüsselenzym der mitochondrialen Atmungskette. Diese Interaktion erhöht das mitochondriale Membranpotenzial, steigert die ATP-Produktion und erzeugt kontrollierte Mengen reaktiver Sauerstoffspezies (ROS), die alle für den Zellstoffwechsel und die Signalübertragung essenziell sind. Klinische Studien haben gezeigt, dass tPBM die kognitive Funktion signifikant verbessern kann, insbesondere bei Menschen mit Schädel-Hirn-Trauma, schweren Depressionen und anderen neurologischen Erkrankungen. Die Fähigkeit der tPBM, die mitochondriale Aktivität und die ROS-Produktion zu modulieren, untermauert ihr Potenzial, die Neuroplastizität zu fördern und die Gehirngesundheit zu unterstützen.
Neuroplastizität verstehen: Die grundlegenden Mechanismen
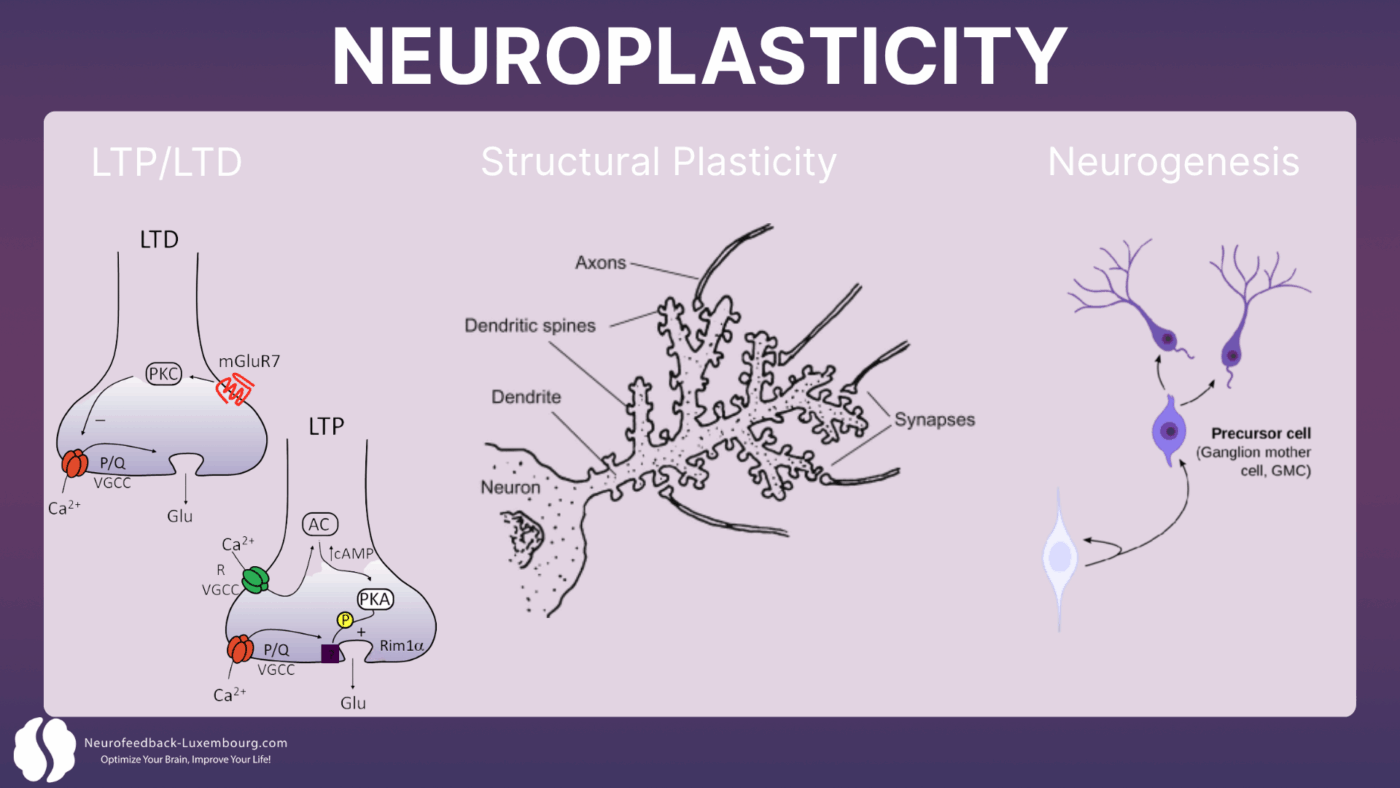
Synaptische Plastizität: Die Grundlage der Anpassung
Neuroplastizität wirkt in erster Linie durch vier miteinander verbundene Mechanismen:
1. Langzeitpotenzierung (LTP)Die dauerhafte Stärkung synaptischer Verbindungen
2. Langzeitdepression (LTD)Die selektive Schwächung ungenutzter Verbindungen
3. Strukturelle PlastizitätDie Bildung neuer dendritischer Dornen und Synapsen
4. NeurogeneseDie Bildung neuer Neuronen in bestimmten Hirnregionen
Diese Prozesse erfordern präzise Synchronisation elektrische Aktivität, erhebliche Energieverfügbarkeit und ein optimales biochemisches Umfeld sind Voraussetzung. Genau hier setzt tPBM als … an neuroplastischer Katalysator.
Lichtenergie und Gehirnfunktion
Lichtenergie, insbesondere im nahen Infrarotbereich, spielt eine entscheidende Rolle bei der Optimierung der Gehirnfunktion. Bei Anwendung mittels Low-Level-Lichttherapie (LLLT) dringt Nahinfrarotlicht in biologisches Gewebe ein und interagiert mit zellulären Bestandteilen, wodurch eine Kaskade positiver Effekte ausgelöst wird (1)(2). Studien haben gezeigt, dass tPBM die Hirndurchblutung steigern und so die Versorgung aktiver Hirnregionen mit Sauerstoff und Nährstoffen verbessern kann (3)(4). Diese verbesserte Perfusion, kombiniert mit der Hochregulierung von Wachstumsfaktoren wie dem vaskulären endothelialen Wachstumsfaktor (VEGF), fördert die Gewebereparatur und -regeneration (5). Darüber hinaus stimuliert tPBM nachweislich die Neurogenese und synaptische Plastizität, die beide für Lernen und Gedächtnis essenziell sind (6). Die Reduktion von oxidativem Stress und Entzündungen trägt zusätzlich zu einem gesunden neuronalen Milieu bei. Für optimale Ergebnisse verwenden tPBM-Protokolle typischerweise Leistungsdichten im Bereich von 10–100 mW/cm² und Energiedosen zwischen 1–10 J/cm², um eine effektive Lichtzufuhr zum Zielgewebe bei gleichzeitig maximaler Sicherheit und Wirksamkeit zu gewährleisten. Diese Parameter sind unerlässlich, um das volle Potenzial der Lichttherapie bei der Verbesserung der Gehirnfunktion und der Unterstützung der Gewebereparatur auszuschöpfen.
Lichtübertragung an das Gehirn
Die effektive Lichtapplikation an das Gehirn ist ein Eckpfeiler einer erfolgreichen tPBM-Therapie. Verschiedene Ansätze wurden entwickelt, um sicherzustellen, dass Nahinfrarotlicht die gewünschten Zielgewebe erreicht. Bei der transkraniellen tPBM wird Infrarotlicht direkt auf die Kopfhaut appliziert, wodurch ein tiefes Eindringen in kortikale und subkortikale Regionen ermöglicht wird. Die intranasale tPBM leitet das Licht durch die Nasenhöhle und ermöglicht so den Zugang zu Hirnstrukturen über das dichte Gefäßnetz in diesem Bereich. Extrakranielle Methoden, wie die Applikation von Licht auf Hals oder Rumpf, können systemische Effekte stimulieren, die indirekt die Hirngesundheit fördern. Die Wahl der Applikationsmethode hängt von der jeweiligen neurologischen Erkrankung und den Therapiezielen ab. Nahinfrarotlicht wird aufgrund seiner überlegenen Gewebepenetration dem sichtbaren Licht vorgezogen, da es tiefer liegende Hirnstrukturen erreicht, die für kürzere Wellenlängen unzugänglich sind (2). Das Verständnis der Prinzipien der Lichtpenetration und der einzigartigen Eigenschaften von Nahinfrarotlicht ist essenziell für die Optimierung der tPBM-Ergebnisse.
Wie tPBM die Neuroplastizität direkt aktiviert
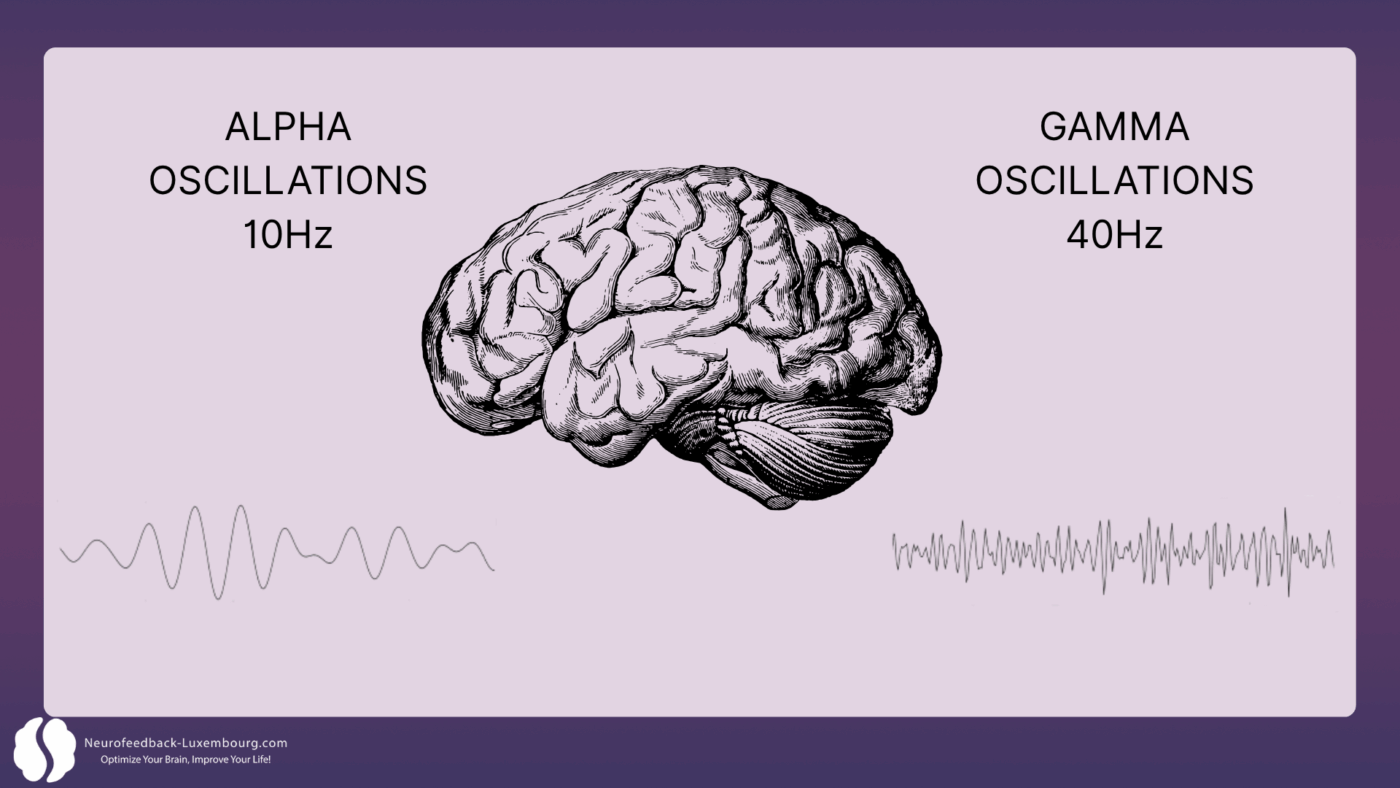
Modulation neuronaler Oszillationen: Der Zeitpunkt der Plastizität
Neuroplastizität hängt grundlegend ab von präzise Zeitmessung Studien belegen, dass tPBM die neuronalen Aktivitätsmuster direkt beeinflusst und so plastische Veränderungen steuert
Gamma-Oszillationen (40 Hz) und LTP:
- Es konnte gezeigt werden, dass eine Stimulation mit 40 Hz das Gedächtnis und das Lernvermögen verbessert und gleichzeitig Amyloid-Plaques reduziert (1). Diese Modulation führt vermutlich zu einer Erhöhung der Anzahl nicht-entzündlicher Mikroglia, die für den Abbau von Amyloid-Plaques verantwortlich sind. Die Zufuhr von gepulster NIR-Lichtenergie mit 40 Hz erhöht die Gamma-Wellenleistung signifikant und reduziert gleichzeitig niedrigere Frequenzen. Dies fördert eine verstärkte Theta-Gamma-Kopplung, die mit verbessertem räumlichem Lernen und gesteigerter kognitiver Leistung korreliert.
- Ergebnis: beschleunigte Stärkung synaptischer Verbindungen während des Lernprozesses.
Alpha-Oszillationen (10 Hz) und Konsolidierung:
- Alpha-Rhythmen (10 Hz) fördern Achtsamkeit, Lernen und Entspannung. Der 10-Hz-Alpha-Modus zeigte in klinischen Studien im Vergleich zu 40 Hz überlegene Ergebnisse mit signifikant besseren kognitiven Resultaten über einen Behandlungszeitraum von 12 Wochen. PBM kann die hippocampale-kortikale Kopplung beeinflussen, die für Lernen und Gedächtnis essenziell ist (1).
- Ergebnisse: Verbessert Lernbegleitung und Stabilisierung entspannter, konzentrierter Zustände.
Vielight Forschungsergebnisse belegen, dass tPBM direkt mitreißen Diese Frequenzen schaffen optimale Zeitfenster für die Herbeiführung plastischer Veränderungen.
Aktivierung von Signalwegen der Plastizität
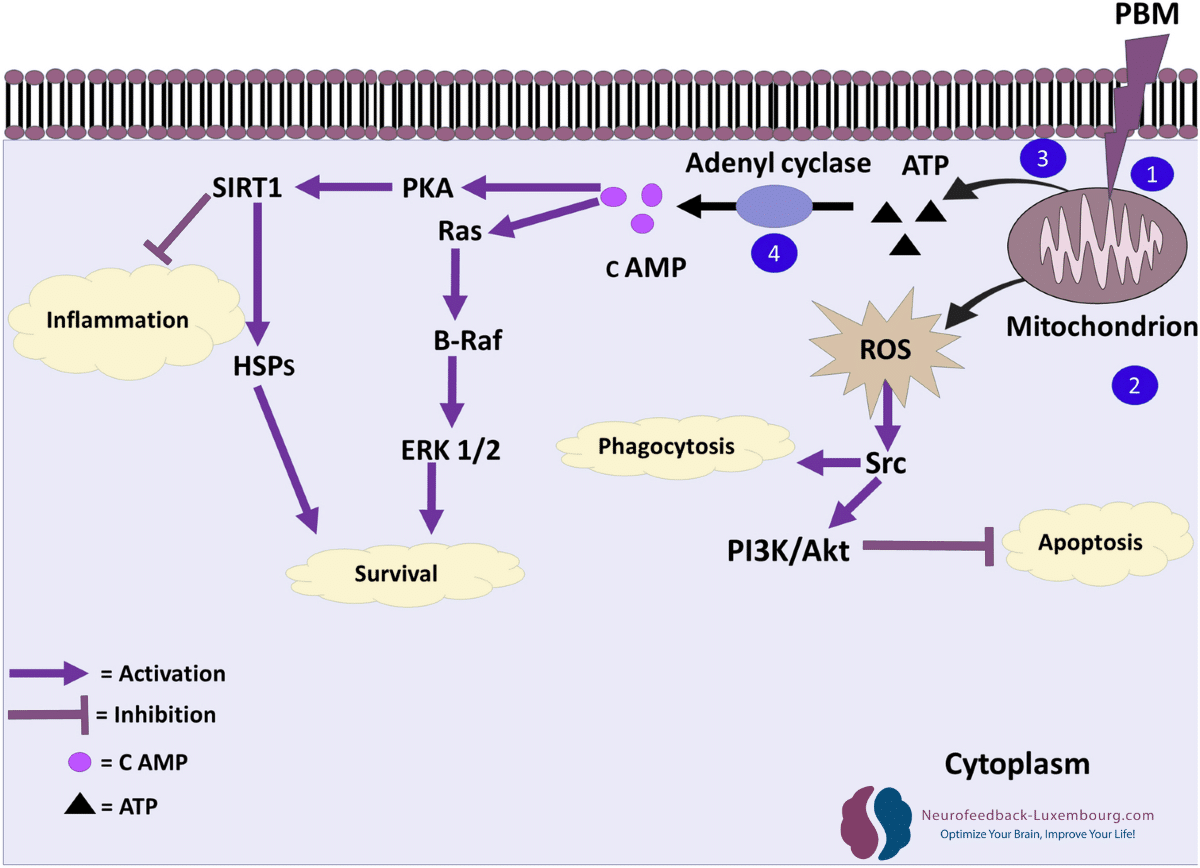
tPBM löst gezielt die molekularen Kaskaden aus, die die Neuroplastizität steuern:
Akt/GSK3β/β-Catenin-Signalweg:
- Direkte Aktivierung durch mitochondriale Stimulation
- Förderung von neuronales Überleben und dendritisches Wachstum
- Erleichterung von Strukturelle Plastizität
CREB (cAMP-Response-Element-Binding)-Signalisierung:
- Erhöhte CREB-Phosphorylierung durch gesteigertes ATP
- Hochregulierung von Plastizitätsgene (Arc, c-Fos, Zif268)(7) (8)
- Beschleunigte Synthese von Proteinen, die für synaptische Veränderungen notwendig sind
tPBM kann auch Transkriptionsfaktoren wie AP-1 und NF-κB beeinflussen, die die Genexpression als Reaktion auf zelluläre Signale regulieren.
mTOR (mechanistisches Ziel von Rapamycin)-Kaskade:
- Aktivierung durch verbesserten Energiestoffwechsel
- Regulierung von lokale Proteinsynthese an Synapsen
- Kontrolle des dendritischen Wachstums und der Dornenbildung (9)
Über diese molekularen Mechanismen hinaus kann tPBM lichtempfindliche Ionenkanäle, einschließlich transienter Rezeptorpotentialkanäle, aktivieren, was zu seinen Auswirkungen auf die neuronale Signalübertragung beiträgt.
BDNF: Der zentrale Regulator der Neuroplastizität
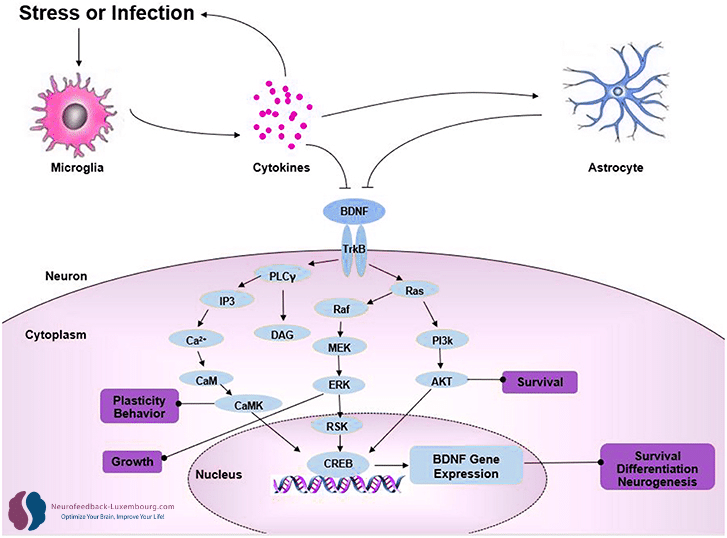
Massive Hochregulierung durch tPBM
Der vom Gehirn stammende neurotrophe Faktor (BDNF) steuert nahezu alle Aspekte der Neuroplastizität. tPBM erzeugt eine dramatischer Anstieg in BDNF-Spiegeln bis (10):
Transkriptionsmechanismen:
- Aktivierung des BDNF-Promotors über phosphoryliertes CREB(11)
- Erhöhte BDNF-mRNA-Expression in Hippocampus und Kortex
- Erhöhte Synthese von Pro-BDNF und reifem BDNF
Direkte funktionelle Effekte:
- Plastizität der WirbelsäuleFörderung der Bildung und Reifung dendritischer Dornen
- Synaptische StärkeErleichterung der AMPA-Rezeptor-Insertion
- Strukturelle PlastizitätStimulation des axonalen und dendritischen Wachstums
BDNF und kritische Periodenplastizität
Forschungsergebnisse zeigen, dass tPBM kann kritische Fenster wieder öffnen der Plastizität bei Erwachsenen (12):
- Reaktivierung juveniler Plastizitätsmechanismen
- Verringerung der perineuronalen Netze, die die Plastizität einschränken
- Wiederherstellung okuläre Dominanzplastizität in Tiermodellen
Diese Fähigkeit legt nahe, dass tPBM die altersabhängige Einschränkungen der Neuroplastizität.
Neurogenese im Erwachsenenalter: Die Entstehung neuer Neuronen
Direkte Stimulation neurogener Nischen
Im Gegensatz zu Ansätzen, die lediglich die bestehende neuronale Aktivität modulieren, stimuliert tPBM die Bildung neuer Neuronen in Tiermodellen:
Subventrikuläre Zone (SVZ) (13):
- Verstärkte Proliferation neuronaler Stammzellen
- Verbesserte Migration in Zielregionen
- Erhöhte Differenzierung in funktionelle Neuronen
Hippocampus Gyrus dentatus:
- Aktivierung von Typ-1-Stammzellen (radiale Gliazellen)
- Förderung der Reifung von Körnerzellen
- Funktionale Integration in bestehende Schaltungen
Molekulare Mechanismen:
- Aktivierung von latentem TGF-β1 zur Förderung der Neurogenese (14)(15)
- Verstärkte Wnt-Signalgebung unterstützt die Stammzellproliferation
- Verminderte entzündungshemmende Wirkung auf die Neurogenese
Funktionelle Auswirkungen der Neurogenese
Durch tPBM neu gebildete Neuronen tragen insbesondere zu Folgendem bei:
- Mustertrennung: Unterscheidung zwischen ähnlichen Erinnerungen (16)
- Kognitive FlexibilitätAnpassung an neue Umgebungen (17)
- StimmungsregulationModulation emotionaler Schaltkreise
- LernfähigkeitVerbesserter Erwerb neuer Fähigkeiten
Synaptogenese: Aufbau neuer Schaltkreise
Beschleunigte Synapsenbildung
tPBM beeinflusst direkt Synaptogenese—die Bildung neuer synaptischer Verbindungen:
Axonales Aussprossen:
- Stimulierung des Wachstumskegelfortschritts
- Verstärkte Expression von Axonleitungsproteinen
- Erleichterung der Bildung neuer synaptischer Boutons
dendritische Dornenbildung:
- Erhöhte Dornendichte in Tierstudien (18)
- Verbesserte Reifung unreifer Stacheln
- Stabilisierung neuer Verbindungen durch koordinierte Aktivitäten
Zusammenbau von synaptischen Komplexen:
- Hochregulierung von Gerüstproteinen (PSD-95, SHANK)
- Verstärkte Clusterbildung von Neurotransmitterrezeptoren
- Verbesserter vesikulärer Transport und verbesserte Neurotransmitterfreisetzung
Schaltungsabhängige Spezifität
Bemerkenswerterweise beeinflusst tPBM die Synaptogenese in einem schaltungsspezifisch Benehmen:
- Verbesserte Konnektivität in Netzwerken aktiv während der Behandlung
- Selektive Stärkung von funktional relevant Verbindungen
- Verbesserte Beschneidung ungenutzter Verbindungen über LTD
Diese Spezifität erklärt, warum tPBM Potentiates Neurofeedback-Effekte ohne zufällige Veränderungen zu erzeugen.
Homöostatische Plastizität: Aufrechterhaltung des Gleichgewichts
Regulation der neuronalen Erregbarkeit
tPBM-Einflüsse homöostatische Plastizität Mechanismen zur Aufrechterhaltung der Netzwerkstabilität:
Synaptische Skalierung:
- Globale Anpassung der synaptischen Stärke
- Prävention von Übererregbarkeit oder Untererregbarkeit
- Aufrechterhaltung optimaler Dynamikbereiche
Intrinsische Plastizität:
- Modulation von spannungsgesteuerten Ionenkanälen
- Anpassung der neuronalen Erregungsschwelle
- Optimierung der dendritischen Integration
Das Selbstregulierung stellt sicher, dass die induzierten plastischen Veränderungen innerhalb gesunder physiologischer Grenzen bleiben.
Die Stiftung für zelluläre Energie: Die Rolle der Cytochrom-c-Oxidase
Erhöhte ATP-Produktion: Treibstoff für die neuronale Anpassung
Neuroplastische Veränderungen sind grundsätzlich energieintensive Prozesse. Die Bildung neuer Synapsen, die dendritische Verzweigung und die Myelinremodellierung erfordern erhebliche zelluläre Energieressourcen. tPBM setzt genau hier an Cytochrom-c-Oxidase-Stimulation—das lichtempfindliche Enzym innerhalb des Komplexes IV der mitochondrialen Elektronentransportkette.
Wenn Nahinfrarotlicht (810 nm) in Hirngewebe eindringt, zielt es gezielt auf dieses mitochondriale Enzym ab und löst eine Kaskade bioenergetischer Effekte aus:
- Sofortiger ATP-AnstiegStudien belegen einen signifikanten Anstieg der Adenosintriphosphat-Produktion innerhalb weniger Minuten nach der Anwendung von tPBM
- Verbesserte oxidative PhosphorylierungEine verbesserte Effizienz der Elektronentransportkette unterstützt eine nachhaltige Energieproduktion
- Optimierung des mitochondrialen MembranpotenzialsWiederherstellung optimaler elektrischer Gradienten über die Mitochondrienmembranen
Das Stoffwechselsteigerung schafft ein ideales zelluläres Umfeld, in dem Neuronen über die energetischen Ressourcen verfügen, um eine schnelle Anpassung und Lernfähigkeit zu unterstützen.
Verbesserte Mikrozirkulation: Die Gefäßstiftung
Neben der direkten Energieproduktion stimuliert tPBM die Stickoxid-(NO)-Synthese und führt so zu messbaren Verbesserungen in folgenden Bereichen:
- zerebrale DurchblutungDie regionale Durchblutung nimmt nach tPBM-Sitzungen zu
- Sauerstoffzufuhr: Erhöhte Hämoglobinsättigung und Gewebeoxygenierung in den behandelten Regionen
- NährstofftransportVerbesserte Zufuhr von Glukose, Aminosäuren und neurotrophen Faktoren zu aktiven neuronalen Netzwerken
Forschungsarbeiten belegen signifikante Steigerungen der Hirndurchblutung in den Frontal-, Temporal- und Okzipitallappen sowie im Hippocampus nach einer tPBM-Behandlung. Diese vaskulären Verbesserungen führen zu einer nährstoffreiche Mikroumgebung das eine anhaltende neuroplastische Aktivität unterstützt.
Entzündungshemmende Neuroprotektion
Kontrollierte Produktion reaktiver Sauerstoffspezies (ROS)
Während übermäßiger oxidativer Stress Nervengewebe schädigt, erzeugt tPBM kontrollierten, milden reaktiven Sauerstoffspezies (ROS) die als nützliche Signalmoleküle fungieren. Diese hormetische Reaktion löst Folgendes aus:
- Hochregulierung antioxidativer EnzymeErhöhte Aktivität von Katalase, Superoxiddismutase und Glutathionperoxidase
- Produktion entzündungshemmender Zytokine: Erhöhte IL-10-Werte und reduzierte proinflammatorische Marker (TNF-α, IL-1β)
- Modulation des Mikroglia-Phänotyps: Wechsel von M1 (entzündlicher) zu M2 (reparativer) Mikroglia-Aktivierung
Das neuroprotektive Umgebung Verhindert Zellschäden, die typischerweise neuroplastische Prozesse unter Stress oder nach Nervenverletzungen behindern. Klinische Integration: Maximierung der Neuroplastizität
Synergistisch tPBM-Neurofeedback-Protokolle
Die optimale Integration nutzt die Zeitfenster der Neuroplastizität:
Vorbereitungsphase (tPBM 20 min):
- Vielight Neuro Gamma (40 Hz): Aktivierung von Aufmerksamkeitsnetzwerken
- Aktivierung von LTP-Mechanismen für nachfolgendes Lernen
- Schnelle Hochregulierung von BDNF und plastischen Faktoren
Trainingsphase (Neurofeedback):
- Anwendung von QEEG-gesteuerten Protokollen während optimaler Kunststofffenster
- Nutzung verbesserter neuronaler Synchronisation
- Nutzung erhöhter kontrollierter Erregbarkeit
Konsolidierungsphase (tPBM 20 min):
- Vielight Neuro Alpha (10 Hz): Förderung der Gedächtniskonsolidierung
- Stabilisierung neu erlernter Muster
- Verbesserte Proteinsynthese unterstützt strukturelle Veränderungen
Neuroplastizitäts-Biomarker
Wir überwachen die Wirksamkeit durch spezifische neuroplastische Indikatoren:
QEEG-Marker:
- Erhöhte Kohärenz in trainierten Frequenzbändern
- Verbesserte Kreuzfrequenzkopplung (Theta-Gamma)
- Verbesserte Kennzahlen zur Netzwerkeffizienz
Verhaltensbeurteilungen:
- Beschleunigte Lernkurven während des Trainings
- Verbesserte Behaltensleistung zwischen den Sitzungen
- Verbesserte Generalisierung auf ungeübte Aufgaben
Fazit: Die neuroplastische Revolution
Die transkranielle Photobiomodulation stellt eine Paradigmenwechsel in unserem Ansatz zur Neuroplastizität. Anstatt einfach anregend das Gehirn verändern, tPBM aktiviert die grundlegenden Mechanismen die alle neuroplastischen Veränderungen ermöglichen.
Diese Technologie ermöglicht uns Folgendes:
- Wichtige Fenster wieder öffnen der Plastizität in jedem Alter
- Die Prozesse beschleunigen der Langzeitpotenzierung (LTP), der Neurogenese und der Synaptogenese
- Optimieren Sie das Timing neuroplastischen Interventionen
- Maximierung der Kundenbindung und Verallgemeinerung von Veränderungen
Für Neurofeedback-Anwender bietet tPBM die einzigartige Möglichkeit, Arbeit mit Neuroplastizität statt gegen seine natürlichen Grenzen anzukämpfen. Das Ergebnis: Veränderungen, die schneller, nachhaltiger und tiefgreifender sind als je zuvor möglich.
Entdecken Sie noch heute Ihr Potenzial zur Gehirnoptimierung. Besuchen Sie unsere Website, um mehr zu erfahren Neurofeedback und unser integrierter Ansatz, und planen Sie Ihren Termin Vorläufige telemedizinische BeratungLassen Sie uns gemeinsam erforschen, wie die transkranielle Photobiomodulation Ihre Gehirngesundheit verändern kann.
Die Zukunft der Gehirnoptimierung liegt im Verständnis und der direkten Aktivierung neuroplastischer Mechanismen. Bei Neurofeedback Luxembourg kombinieren wir QEEG-Expertise mit der Vielight-Technologie, um Gehirntrainingsprogramme zu entwickeln, die … neurologisch informiert Und neuroplastisch optimiert.
Alle Protokolle dienen der Optimierung des Wohlbefindens und stellen keine medizinische Behandlung dar. Die Ergebnisse können individuell variieren. Für eine individuelle Beurteilung wird eine Beratung empfohlen.
Referenzen
(2) Naeser, MA, Martin, PI, Ho, MD, Krengel, MH, Bogdanova, Y., Knight, JA, Hamblin, MR, Fedoruk, AE, Poole, LG, Cheng, C., & Koo, B. (sd). Transkranielle Photobiomodulationstherapie: Signifikante Verbesserungen bei vier ehemaligen Footballspielern mit möglicher chronisch-traumatischer Enzephalopathie. Journal of Alzheimer's Disease Reports, 7(1), 77‑105. https://doi.org/10.3233/ADR-220022
(1) Lim, L. (2024). Modifizierung der Pathophysiologie der Alzheimer-Krankheit durch Photobiomodulation: Modell, Evidenz und Zukunft mit EEG-gesteuerter Intervention. Frontiers in Neurology, 15. https://doi.org/10.3389/fneur.2024.1407785
(3) Chao, LL (2019). Auswirkungen von Photobiomodulationsbehandlungen zu Hause auf die kognitive und Verhaltensfunktion, die zerebrale Perfusion und die funktionelle Konnektivität im Ruhezustand bei Patienten mit Demenz: Eine Pilotstudie. Photobiomodulation, Photomedizin und Laserchirurgie, 37(3), 133‑141. https://doi.org/10.1089/photob.2018.4555
(4) Chao, LL, Barlow, C., Karimpoor, M., & Lim, L. (2020). Veränderungen der Hirnfunktion und -struktur nach selbstverabreichter Photobiomodulationsbehandlung zu Hause bei einem Patienten mit Gehirnerschütterung. Frontiers in Neurology, 11. https://doi.org/10.3389/fneur.2020.00952
(5) Di Gregorio, E., Staelens, M., Hosseinkhah, N., Karimpoor, M., Liburd, J., Lim, L., Shankar, K., & Tuszyński, JA (2024). Raman-Spektroskopie enthüllt photobiomodulationsinduzierten α-Helix-zu-β-Faltblatt-Übergang in Tubulinen: Mögliche Implikationen für Alzheimer und andere neurodegenerative Erkrankungen. Nanomaterialien, 14(13), Artikel 13. https://doi.org/10.3390/nano14131093
(6) Lim, L., Hosseinkhah, N., Van Buskirk, M., Berk, A., Loheswaran, G., Abbaspour, Z., Karimpoor, M., Smith, A., Ho, KF, Pushparaj, A., Zahavi, M., White, A., Rubine, J., Zidel, B., Henderson, C., Clayton, RG, Tingley, DR, Miller, DJ, Karimpoor, M. & Hamblin, MR (2024). Photobiomodulationstherapie mit einem Heimgerät zur Behandlung von COVID-19: Eine randomisierte, kontrollierte Studie zur Wirksamkeit und Sicherheit. Photobiomodulation, Photomedizin und Laserchirurgie, 42(6), 393‑403. https://doi.org/10.1089/pho.2023.0179
(7) Knapska, E., & Kaczmarek, L. (2004). Ein Gen für neuronale Plastizität im Säugetiergehirn: Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Fortschritte in der Neurobiologie, 74(4), 183‑211. https://doi.org/10.1016/j.pneurobio.2004.05.007
(8) Penke, Z., Morice, E., Veyrac, A., Gros, A., Chagneau, C., LeBlanc, P., Samson, N., Baumgärtel, K., Mansuy, IM, Davis, S., & Laroche, S. (2014). Funktionsgewinn von Zif268/Egr1 erleichtert die synaptische Plastizität im Hippocampus und das räumliche Langzeitgedächtnis. Philosophical Transactions of the Royal Society B: Biologische Wissenschaften, 369(1633), 20130159. https://doi.org/10.1098/rstb.2013.0159
(9) Lee, DY (2015). Die Rolle der mTOR-Signalübertragung in der Gehirnentwicklung. Experimentelle Neurobiologie, 24(3), 177‑185. https://doi.org/10.5607/en.2015.24.3.177
(11) Heo, J.-C., Park, J.-A., Kim, D.-K., & Lee, J.-H. (2019). Photobiomodulation (660 nm)-Therapie reduziert oxidativen Stress und induziert die BDNF-Expression im Hippocampus. Wissenschaftliche Berichte, 9(1), 10114. https://doi.org/10.1038/s41598-019-46490-4
(10) de Oliveira, BH, Lins, EF, Kunde, NF, Salgado, ASI, Martins, LM, Bobinski, F., Vieira, WF, Cassano, P., Quialheiro, A., & Martins, DF (2024). Transkranielle Photobiomodulation erhöht die Kognition und den Serum-BDNF-Spiegel bei Erwachsenen über 50 Jahren: Eine randomisierte, doppelblinde, placebokontrollierte Studie. Zeitschrift für Photochemie und Photobiologie B: Biologie, 260, 113041. https://doi.org/10.1016/j.jphotobiol.2024.113041
(12) Hensch, TK, & Bilimoria, PM (2012). Re-opening Windows : Manipulating Critical Periods for Brain Development. Cerebrum: Das Dana-Forum für Hirnforschung, 2012, 11.
(13) Chang, S.-Y., & Lee, MY (2023). Photobiomodulation der Neurogenese durch Steigerung der Differenzierung von Stammzellen und neuronalen Vorläuferzellen im zentralen und peripheren Nervensystem. Internationale Zeitschrift für Molekularwissenschaften, 24(20), 15427. https://doi.org/10.3390/ijms242015427
(14) Wu, X., Shen, Q., Zhang, Z., Zhang, D., Gu, Y., & Xing, D. (2021). Photoaktivierung des TGFβ/SMAD-Signalwegs verbessert die adulte hippocampale Neurogenese im Alzheimer-Krankheitsmodell. Stammzellenforschung und -therapie, 12(1), 345. https://doi.org/10.1186/s13287-021-02399-2
(15) Kandasamy, M., Lehner, B., Kraus, S., Sander, PR, Marschallinger, J., Rivera, FJ, Trümbach, D., Ueberham, U., Reitsamer, HA, Strauss, O., Bogdahn, U., Couillard-Despres, S., & Aigner, L. (2014). TGF-beta-Signalgebung in der adulten neurogenen Nische fördert sowohl die Stammzellruhe als auch die Bildung neuer Neuronen. Zeitschrift für Zell- und Molekularmedizin, 18(7), 1444‑1459. https://doi.org/10.1111/jcmm.12298
(16) Johnston, ST, Shtrahman, M., Parylak, S., Gonçalves, JT, & Gage, FH (2016). Paradox der Musterseparation und der adulten Neurogenese: Eine Doppelrolle für neue Neuronen beim Ausgleich von Gedächtnisauflösung und Robustheit. Neurobiologie des Lernens und des Gedächtnisses, 129, 60‑68. https://doi.org/10.1016/j.nlm.2015.10.013
(17) Anacker, C., & Hen, R. (2017). Neurogenese im Hippocampus Erwachsener und kognitive Flexibilität – Verknüpfung von Gedächtnis und Stimmung. Nature Reviews Neurowissenschaften, 18(6), 335‑346. https://doi.org/10.1038/nrn.2017.45
(18) Meng, C., He, Z., & Xing, D. (2013). Low-Level-Lasertherapie rettet Dendritenatrophie durch Hochregulierung der BDNF-Expression: Implikationen für die Alzheimer-Krankheit. Das Journal of Neuroscience: Das offizielle Organ der Gesellschaft für Neurowissenschaften, 33(33), 13505‑13517. https://doi.org/10.1523/JNEUROSCI.0918-13.2013
