Neuroplasticity—the brain’s extraordinary capacity to reorganize, modify its connections, and create new neural circuits—constitutes the fundamental mechanism underlying all learning, recovery, and cognitive optimization. At Neurofeedback Luxembourg, we understand that maximizing neuroplasticity requires more than training; it demands the creation of an optimal cellular environment. Transcranial Photobiomodulation (tPBM) through Vielight technology offers a revolutionary approach to activate and amplify neuroplastic mechanisms at their source.
tPBM utilizes non ionizing light sources from the electromagnetic spectrum to interact with the human body at the cellular level.
Introduction to Neuroplasticity and tPBM
Neuroplasticity is the remarkable ability of the brain to reorganize itself by forming new neural connections throughout life. This adaptive process enables the brain to respond to new experiences, recover from injuries, and enhance learning and memory. Transcranial photobiomodulation (tPBM) is an innovative, non-invasive technique that harnesses low level light therapy—including low level laser therapy and light emitting diodes—to stimulate the brain’s natural capacity for change. By delivering near infrared light to the scalp, tPBM targets cytochrome c oxidase, a key enzyme in the mitochondrial respiratory chain. This interaction boosts mitochondrial membrane potential, increases ATP production, and generates controlled levels of reactive oxygen species (ROS), all of which are essential for cellular metabolism and signaling. Clinical studies have shown that tPBM can significantly improve cognitive function, particularly in individuals with traumatic brain injury, major depressive disorder, and other neurological conditions. The ability of tPBM to modulate mitochondrial activity and ROS production underpins its potential to drive neuroplasticity and support brain health.
Understanding Neuroplasticity: The Fundamental Mechanisms
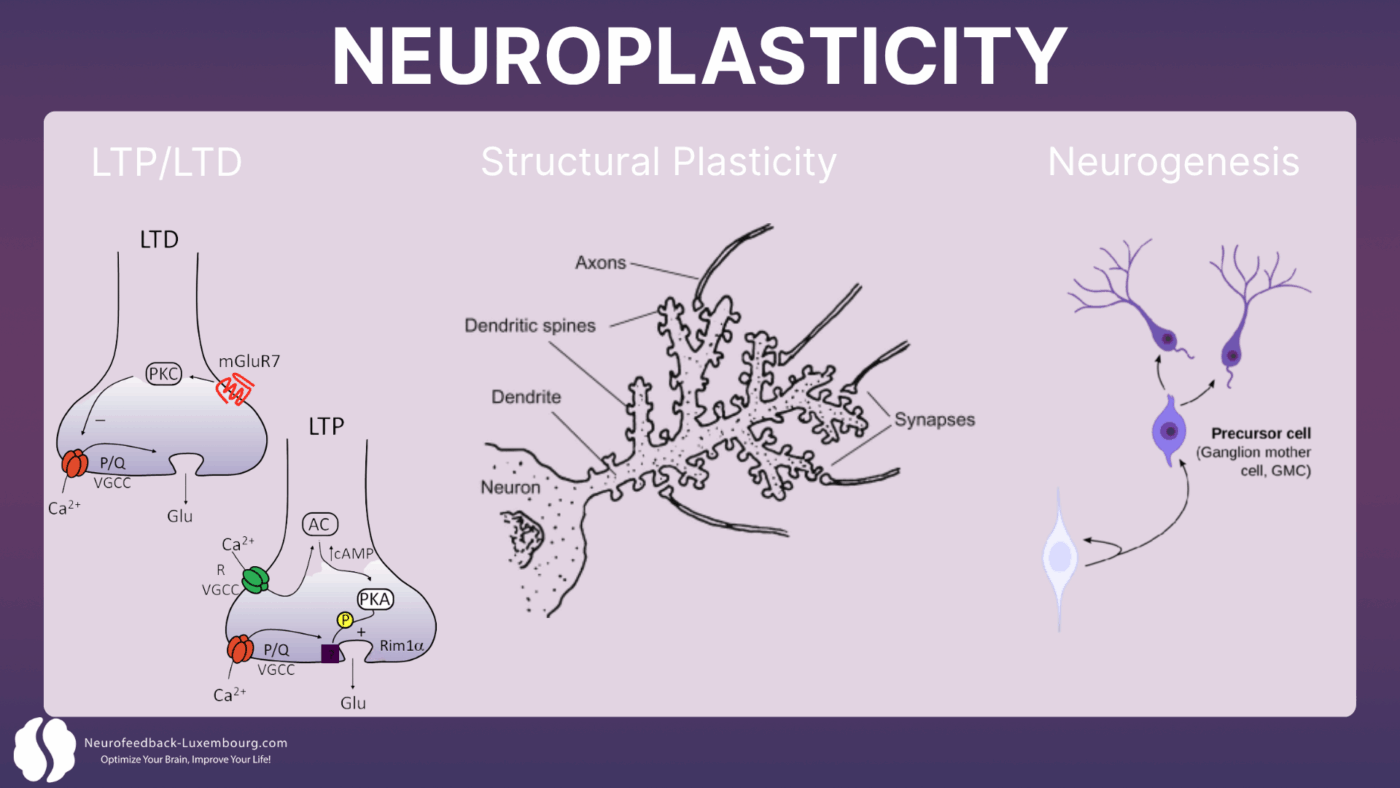
Synaptic Plasticity: The Foundation of Adaptation
Neuroplasticity operates primarily through four interconnected mechanisms:
1. Long-Term Potentiation (LTP): The durable strengthening of synaptic connections
2. Long-Term Depression (LTD): The selective weakening of unused connections
3. Structural plasticity: The formation of new dendritic spines and synapses
4. Neurogenesis: The creation of new neurons in specific brain regions
These processes require precise synchronization of electrical activity, substantial energy availability, and an optimal biochemical environment. This is precisely where tPBM intervenes as a neuroplastic catalyst.
Light Energy and Brain Function
Light energy, especially in the near infrared spectrum, plays a pivotal role in optimizing brain function. When applied through low level light therapy, near infrared light penetrates biological tissue and interacts with cellular components to trigger a cascade of beneficial effects (1)(2). Research has demonstrated that tPBM can increase cerebral blood flow, supporting the delivery of oxygen and nutrients to active brain regions (3)(4). This enhanced perfusion, combined with the upregulation of growth factors such as vascular endothelial growth factor (VEGF), promotes tissue repair and regeneration (5). Additionally, tPBM has been shown to stimulate neurogenesis and synaptic plasticity, both of which are critical for learning and memory (6). The reduction of oxidative stress and inflammation further supports a healthy neural environment. For optimal results, tPBM protocols typically use power densities in the range of 10-100 mW/cm² and energy doses between 1-10 J/cm², ensuring effective light delivery to target tissues while maximizing safety and efficacy. These parameters are essential for harnessing the full potential of light therapy in enhancing brain function and supporting tissue repair.
Delivery of Light to the Brain
Effective delivery of light to the brain is a cornerstone of successful tPBM therapy. Several approaches have been developed to ensure that near infrared light reaches the intended target tissues. Transcranial tPBM involves applying infrared light directly to the scalp, allowing for deep light penetration into cortical and subcortical regions. Intranasal tPBM delivers light through the nasal cavity, providing access to brain structures via the rich vascular network in this area. Extracranial methods, such as applying light to the neck or torso, can stimulate systemic effects that indirectly benefit brain health. The choice of delivery method depends on the specific neurological condition and therapeutic goals. Importantly, near infrared light is favored over visible light due to its superior tissue penetration, enabling it to reach deeper brain structures that are inaccessible to shorter wavelengths (2). Understanding the principles of light penetration and the unique properties of near infrared light is essential for optimizing tPBM outcomes.
How tPBM Directly Activates Neuroplasticity
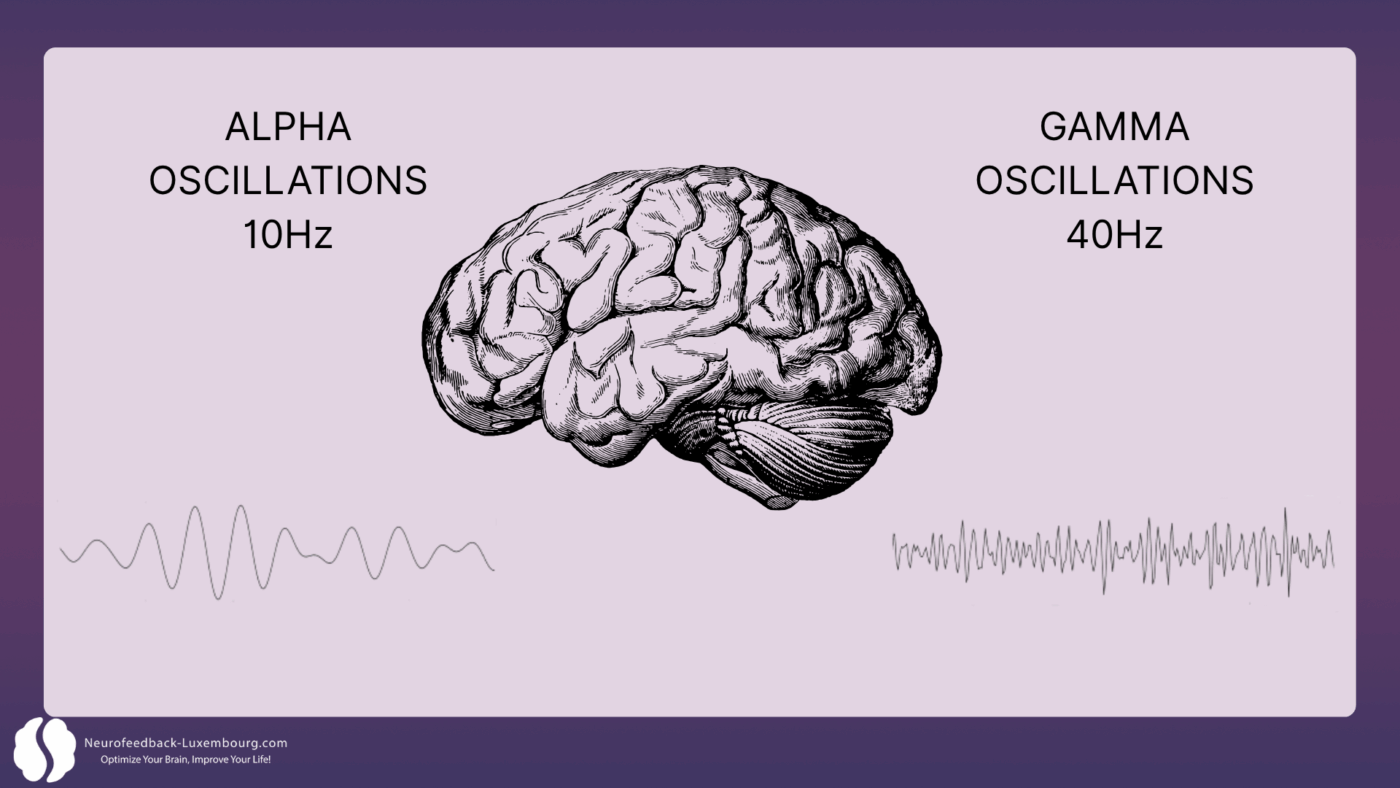
Neural Oscillation Modulation: The Timing of Plasticity
Neuroplasticity fundamentally depends on the precise timing of neuronal activity. Studies demonstrate that tPBM directly influences brain oscillations that orchestrate plastic changes:
Gamma Oscillations (40Hz) and LTP:
- 40Hz stimulation has been shown to improve memory and learning, alongside reduction in amyloid plaques (1). This modulation is hypothesized to increase non-inflammatory microglia, which are responsible for removing amyloid plaque buildup. The delivery of NIR light energy pulsed at 40Hz significantly increases gamma wave power while reducing slower frequencies, facilitating enhanced theta-gamma coupling that correlates with improved spatial learning and cognitive task performance.
- Result: accelerated strengthening of synaptic connections during learning.
Alpha Oscillations (10Hz) and Consolidation:
- Alpha rhythms (10Hz) support mindfulness, learning, and relaxation. The 10Hz Alpha mode has demonstrated superior results compared to 40Hz in clinical studies, with significantly better cognitive outcomes over 12 weeks of treatment. PBM can influence hippocampal-cortical coupling, which is essential for learning and memory (1).
- Results: Enhanced learning facilitation and stabilization of relaxed, focused states.
Vielight research demonstrates that tPBM directly entrains these frequencies, creating optimal temporal windows for inducing plastic changes.
Activation of Plastic Signaling Pathways
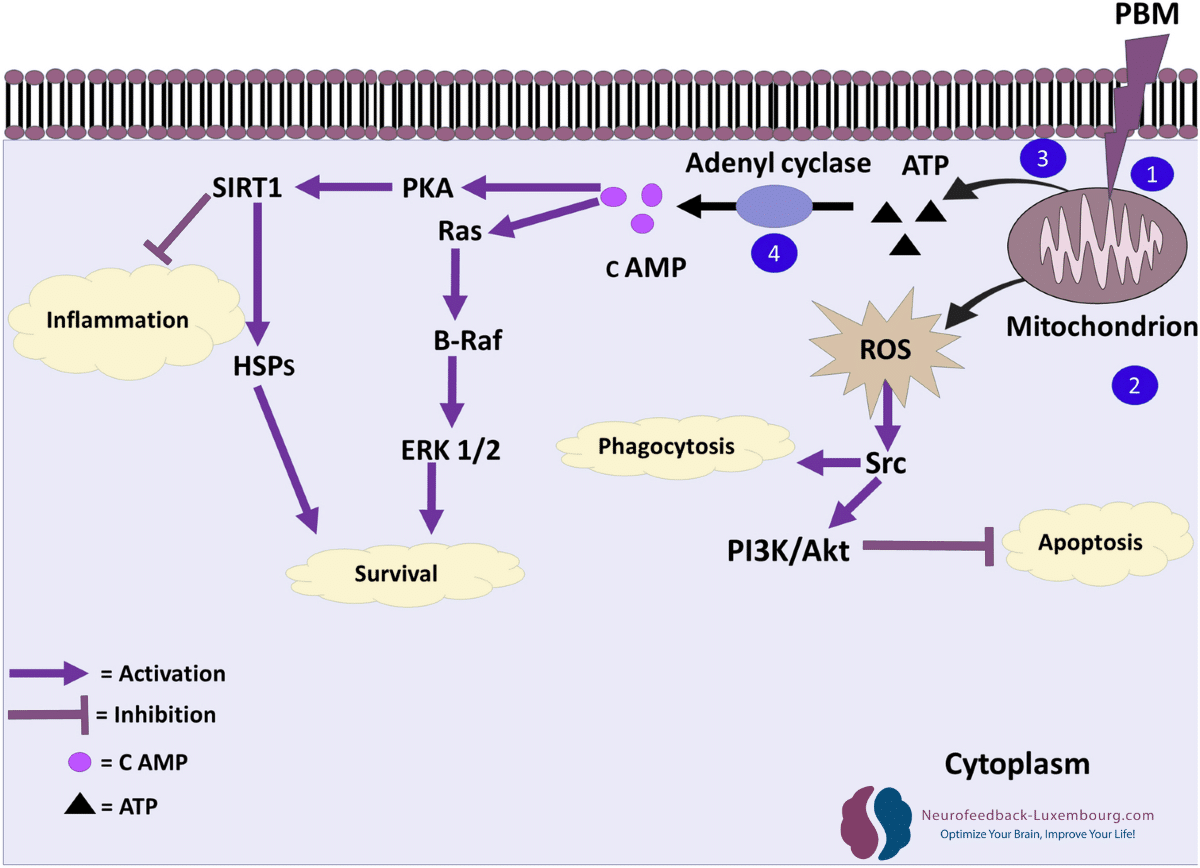
tPBM specifically triggers the molecular cascades that govern neuroplasticity:
Akt/GSK3β/β-catenin Pathway:
- Direct activation through mitochondrial stimulation
- Promotion of neuronal survival and dendritic growth
- Facilitation of structural plasticity
CREB (cAMP Response Element-Binding) Signaling:
- Enhanced CREB phosphorylation through increased ATP
- Upregulation of plasticity genes (Arc, c-Fos, Zif268)(7) (8)
- Accelerated synthesis of proteins necessary for synaptic changes
tPBM can also influence transcription factors such as AP-1 and NF-κB, which regulate gene expression in response to cellular signals.
mTOR (mechanistic Target of Rapamycin) Cascade:
- Activation through improved energy metabolism
- Regulation of local protein synthesis at synapses
- Control of dendritic growth and spine formation (9)
Beyond these molecular mechanisms, tPBM may activate light sensitive ion channels, including transient receptor potential channels, which contribute to its effects on neuronal signaling.
BDNF: The Master Regulator of Neuroplasticity
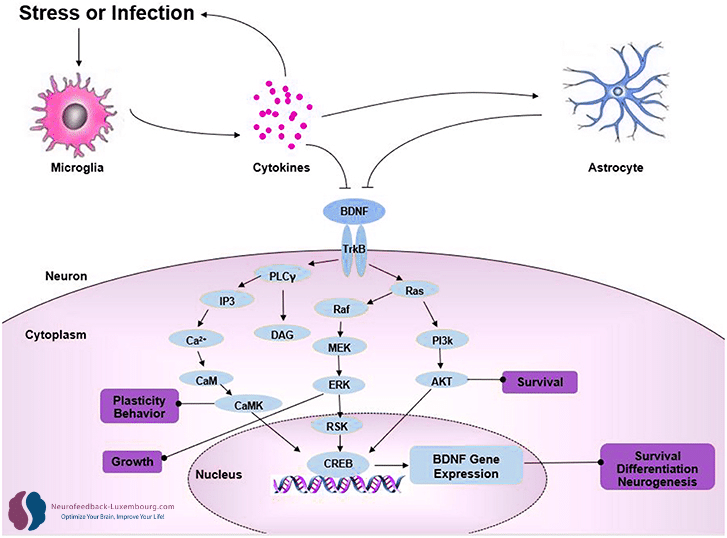
Massive Upregulation by tPBM
Brain-Derived Neurotrophic Factor (BDNF) orchestrates virtually all aspects of neuroplasticity. tPBM produces a dramatic increase in BDNF levels through (10):
Transcriptional mechanisms:
- Activation of BDNF promoter via phosphorylated CREB(11)
- Enhanced BDNF mRNA expression in hippocampus and cortex
- Increased synthesis of pro-BDNF and mature BDNF
Direct functional effects:
- Spine plasticity: Promotion of dendritic spine formation and maturation
- Synaptic strength: Facilitation of AMPA receptor insertion
- Structural plasticity: Stimulation of axonal and dendritic growth
BDNF and Critical Period Plasticity
Research reveals that tPBM can reopen critical windows of plasticity in adults (12):
- Reactivation of juvenile plasticity mechanisms
- Reduction of perineuronal nets limiting plasticity
- Restoration of ocular dominance plasticity in animal models
This capacity suggests that tPBM can overcome the age-dependent limitations of neuroplasticity.
Adult Neurogenesis: Creating New Neurons
Direct Stimulation of Neurogenic Niches
Unlike approaches that merely modulate existing neuronal activity, tPBM stimulates the creation of new neurons in animal models:
Subventricular Zone (SVZ) (13):
- Enhanced proliferation of neural stem cells
- Improved migration to target regions
- Increased differentiation into functional neurons
Hippocampal Dentate Gyrus:
- Activation of type-1 stem cells (radial glia-like)
- Promotion of granule neuron maturation
- Functional integration into existing circuits
Molecular mechanisms:
- Activation of latent TGF-β1 promoting neurogenesis (14)(15)
- Enhanced Wnt signaling supporting stem cell proliferation
- Reduced inflammatory inhibition of neurogenesis
Functional Impact of Neurogenesis
New neurons generated through tPBM specifically contribute to:
- Pattern separation: Discrimination between similar memories (16)
- Cognitive flexibility: Adaptation to new environments (17)
- Mood regulation: Modulation of emotional circuits
- Learning capacity: Enhanced acquisition of new skills
Synaptogenesis: Building New Circuits
Accelerated Synapse Formation
tPBM directly influences synaptogenesis—the formation of new synaptic connections:
Axonal sprouting:
- Stimulation of growth cone advancement
- Enhanced expression of axon guidance proteins
- Facilitation of new synaptic bouton formation
Dendritic spine formation:
- Increased spine density in animal studies (18)
- Enhanced maturation of immature spines
- Stabilization of new connections through coordinated activity
Synaptic complex assembly:
- Upregulation of scaffolding proteins (PSD-95, SHANK)
- Enhanced clustering of neurotransmitter receptors
- Improved vesicular trafficking and neurotransmitter release
Circuit-Dependent Specificity
Remarkably, tPBM influences synaptogenesis in a circuit-specific manner:
- Enhanced connectivity in networks active during treatment
- Selective strengthening of functionally relevant connections
- Improved pruning of unused connections via LTD
This specificity explains why tPBM potentiates neurofeedback effects without creating random changes.
Homeostatic Plasticity: Maintaining Balance
Regulation of Neuronal Excitability
tPBM influences homeostatic plasticity mechanisms that maintain network stability:
Synaptic scaling:
- Global adjustment of synaptic strength
- Prevention of hyperexcitability or hypoexcitability
- Maintenance of optimal dynamic ranges
Intrinsic plasticity:
- Modulation of voltage-gated ion channels
- Adjustment of neuronal excitation threshold
- Optimization of dendritic integration
This self-regulation ensures that induced plastic changes remain within healthy physiological limits.
The Cellular Energy Foundation: Role of Cytochrome c Oxidase
Enhanced ATP Production: Fueling Neural Adaptation
Neuroplastic changes are fundamentally energy-intensive processes. The formation of new synapses, dendritic branching, and myelin remodeling require substantial cellular energy resources. tPBM directly addresses this need through cytochrome c oxidase stimulation—the photosensitive enzyme within Complex IV of the mitochondrial electron transport chain.
When near-infrared light (810nm) penetrates brain tissue, it specifically targets this mitochondrial enzyme, triggering a cascade of bioenergetic effects:
- Immediate ATP surge: Studies demonstrate significant increases in adenosine triphosphate production within minutes of tPBM application
- Enhanced oxidative phosphorylation: Improved efficiency of the electron transport chain supports sustained energy production
- Mitochondrial membrane potential optimization: Restoration of optimal electrical gradients across mitochondrial membranes
This metabolic enhancement creates an ideal cellular environment where neurons possess the energetic resources to support rapid adaptation and learning.
Improved Microcirculation: The Vascular Foundation
Beyond direct energy production, tPBM stimulates nitric oxide (NO) synthesis, producing measurable improvements in:
- Cerebral blood flow: Regional perfusion increases following tPBM sessions
- Oxygen delivery: Enhanced hemoglobin saturation and tissue oxygenation in treated regions
- Nutrient transport: Improved delivery of glucose, amino acids, and neurotrophic factors to active neural networks
Research demonstrates significant increases in cerebral perfusion within the frontal, temporal, and occipital lobes, as well as the hippocampus, following tPBM treatment. These vascular improvements create a nutrient-rich microenvironment that supports sustained neuroplastic activity.
Anti-Inflammatory Neuroprotection
Controlled Reactive Oxygen Species (ROS) Production
While excessive oxidative stress damages neural tissue, tPBM produces controlled, mild reactive oxygen species (ROS) that function as beneficial signaling molecules. This hormetic response triggers:
- Antioxidant enzyme upregulation: Enhanced catalase, superoxide dismutase, and glutathione peroxidase activity
- Anti-inflammatory cytokine production: Increased IL-10 and reduced pro-inflammatory markers (TNF-α, IL-1β)
- Microglial phenotype modulation: Shift from M1 (inflammatory) to M2 (reparative) microglial activation
This neuroprotective environment prevents the cellular damage that typically impedes neuroplastic processes during stress or after neural injury.Clinical Integration: Maximizing Neuroplasticity
Synergistic tPBM-Neurofeedback Protocols
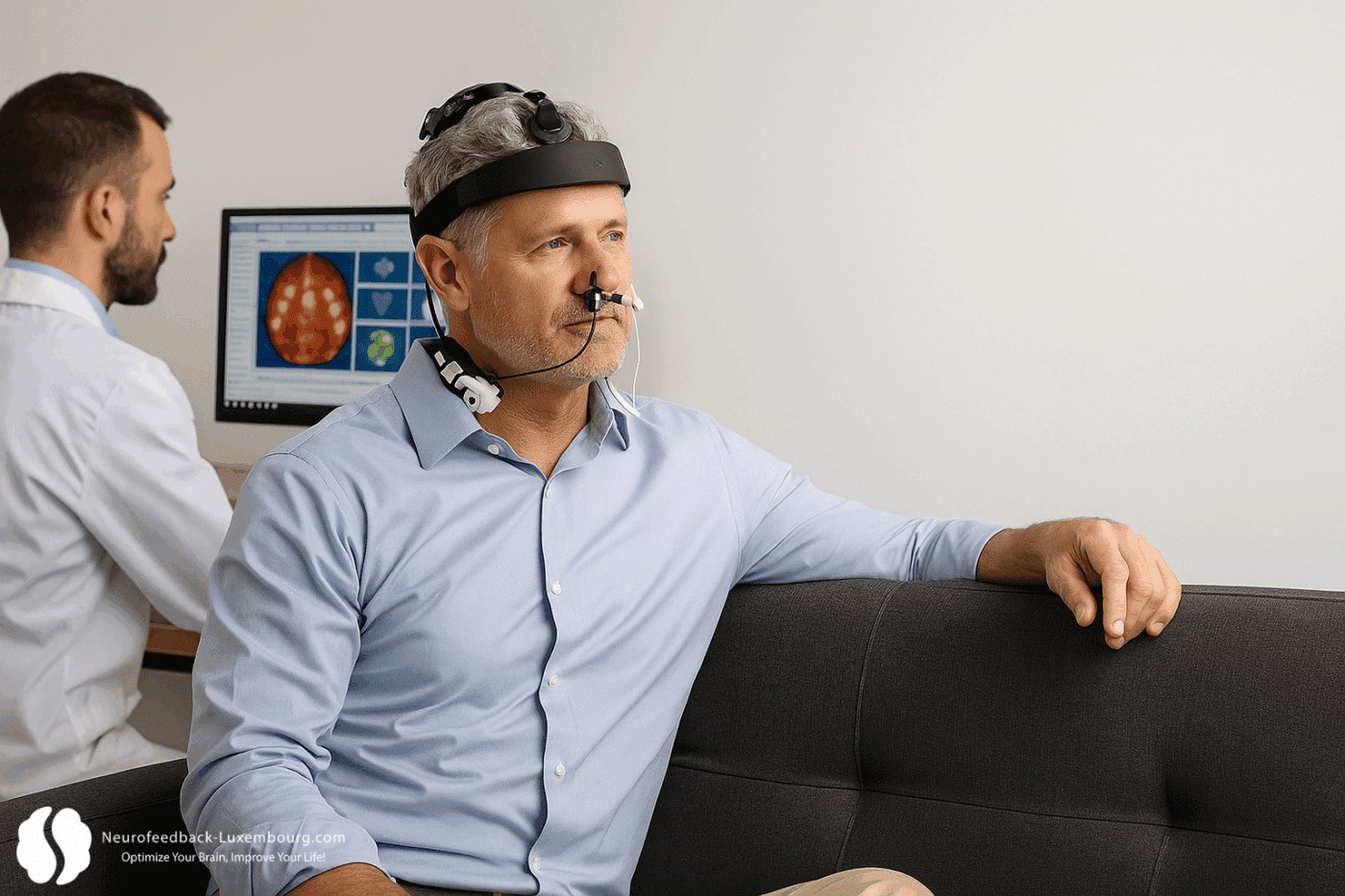
Optimal integration exploits the temporal windows of neuroplasticity:
Preparation Phase (tPBM 20 min):
- Vielight Neuro Gamma (40Hz): Activation of attentional networks
- Priming of LTP mechanisms for subsequent learning
- Rapid upregulation of BDNF and plastic factors
Training Phase (Neurofeedback):
- Application of QEEG-guided protocols during optimal plastic window
- Utilization of enhanced neural synchronization
- Exploitation of increased controlled excitability
Consolidation Phase (tPBM 20 min):
- Vielight Neuro Alpha (10Hz): Facilitation of memory consolidation
- Stabilization of newly learned patterns
- Enhanced protein synthesis supporting structural changes
Neuroplasticity Biomarkers
We monitor efficacy through specific neuroplastic indicators:
QEEG markers:
- Increased coherence in trained frequency bands
- Enhanced cross-frequency coupling (theta-gamma)
- Improved network efficiency metrics
Behavioral assessments:
- Accelerated acquisition curves during training
- Enhanced retention between sessions
- Improved generalization to untrained tasks
Conclusion: The Neuroplastic Revolution
Transcranial Photobiomodulation represents a paradigmatic shift in our approach to neuroplasticity. Rather than simply stimulating the brain to change, tPBM activates the fundamental mechanisms that make all neuroplastic change possible.
This technology enables us to:
- Reopen critical windows of plasticity at any age
- Accelerate the processes of LTP, neurogenesis, and synaptogenesis
- Optimize the timing of neuroplastic interventions
- Maximize retention and generalization of changes
For neurofeedback practitioners, tPBM offers the unique opportunity to work with neuroplasticity rather than against its natural limitations. The result: changes that are faster, more durable, and more profound than ever before possible.
Discover your brain optimization potential today. Visit ou website to learn more about neurofeedback and our integrated approach, and schedule your preliminary teleconsultation. Together, let’s explore how transcranial photobiomodulation can transform your brain health.
The future of brain optimization lies in understanding and directly activating neuroplastic mechanisms. At Neurofeedback Luxembourg, we combine QEEG expertise with Vielight technology to create brain training programs that are neurologically informed and neuroplastically optimized.
All protocols are designed for wellness optimization and do not constitute medical treatment. Individual results may vary. Consultation recommended for personalized assessment.
References
(2) Naeser, M. A., Martin, P. I., Ho, M. D., Krengel, M. H., Bogdanova, Y., Knight, J. A., Hamblin, M. R., Fedoruk, A. E., Poole, L. G., Cheng, C., & Koo, B. (s. d.). Transcranial Photobiomodulation Treatment : Significant Improvements in Four Ex-Football Players with Possible Chronic Traumatic Encephalopathy. Journal of Alzheimer’s Disease Reports, 7(1), 77‑105. https://doi.org/10.3233/ADR-220022
(1) Lim, L. (2024). Modifying Alzheimer’s disease pathophysiology with photobiomodulation : Model, evidence, and future with EEG-guided intervention. Frontiers in Neurology, 15. https://doi.org/10.3389/fneur.2024.1407785
(3) Chao, L. L. (2019). Effects of Home Photobiomodulation Treatments on Cognitive and Behavioral Function, Cerebral Perfusion, and Resting-State Functional Connectivity in Patients with Dementia : A Pilot Trial. Photobiomodulation, Photomedicine, and Laser Surgery, 37(3), 133‑141. https://doi.org/10.1089/photob.2018.4555
(4) Chao, L. L., Barlow, C., Karimpoor, M., & Lim, L. (2020). Changes in Brain Function and Structure After Self-Administered Home Photobiomodulation Treatment in a Concussion Case. Frontiers in Neurology, 11. https://doi.org/10.3389/fneur.2020.00952
(5) Di Gregorio, E., Staelens, M., Hosseinkhah, N., Karimpoor, M., Liburd, J., Lim, L., Shankar, K., & Tuszyński, J. A. (2024). Raman Spectroscopy Reveals Photobiomodulation-Induced α-Helix to β-Sheet Transition in Tubulins : Potential Implications for Alzheimer’s and Other Neurodegenerative Diseases. Nanomaterials, 14(13), Article 13. https://doi.org/10.3390/nano14131093
(6) Lim, L., Hosseinkhah, N., Van Buskirk, M., Berk, A., Loheswaran, G., Abbaspour, Z., Karimpoor, M., Smith, A., Ho, K. F., Pushparaj, A., Zahavi, M., White, A., Rubine, J., Zidel, B., Henderson, C., Clayton, R. G., Tingley, D. R., Miller, D. J., Karimpoor, M., & Hamblin, M. R. (2024). Photobiomodulation Treatment with a Home-Use Device for COVID-19 : A Randomized Controlled Trial for Efficacy and Safety. Photobiomodulation, Photomedicine, and Laser Surgery, 42(6), 393‑403. https://doi.org/10.1089/pho.2023.0179
(7) Knapska, E., & Kaczmarek, L. (2004). A gene for neuronal plasticity in the mammalian brain : Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Progress in Neurobiology, 74(4), 183‑211. https://doi.org/10.1016/j.pneurobio.2004.05.007
(8) Penke, Z., Morice, E., Veyrac, A., Gros, A., Chagneau, C., LeBlanc, P., Samson, N., Baumgärtel, K., Mansuy, I. M., Davis, S., & Laroche, S. (2014). Zif268/Egr1 gain of function facilitates hippocampal synaptic plasticity and long-term spatial recognition memory. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1633), 20130159. https://doi.org/10.1098/rstb.2013.0159
(9) Lee, D. Y. (2015). Roles of mTOR Signaling in Brain Development. Experimental Neurobiology, 24(3), 177‑185. https://doi.org/10.5607/en.2015.24.3.177
(11) Heo, J.-C., Park, J.-A., Kim, D.-K., & Lee, J.-H. (2019). Photobiomodulation (660 nm) therapy reduces oxidative stress and induces BDNF expression in the hippocampus. Scientific Reports, 9(1), 10114. https://doi.org/10.1038/s41598-019-46490-4
(10) de Oliveira, B. H., Lins, E. F., Kunde, N. F., Salgado, A. S. I., Martins, L. M., Bobinski, F., Vieira, W. F., Cassano, P., Quialheiro, A., & Martins, D. F. (2024). Transcranial photobiomodulation increases cognition and serum BDNF levels in adults over 50 years : A randomized, double-blind, placebo-controlled trial. Journal of Photochemistry and Photobiology B: Biology, 260, 113041. https://doi.org/10.1016/j.jphotobiol.2024.113041
(12) Hensch, T. K., & Bilimoria, P. M. (2012). Re-opening Windows : Manipulating Critical Periods for Brain Development. Cerebrum: the Dana Forum on Brain Science, 2012, 11.
(13) Chang, S.-Y., & Lee, M. Y. (2023). Photobiomodulation of Neurogenesis through the Enhancement of Stem Cell and Neural Progenitor Differentiation in the Central and Peripheral Nervous Systems. International Journal of Molecular Sciences, 24(20), 15427. https://doi.org/10.3390/ijms242015427
(14) Wu, X., Shen, Q., Zhang, Z., Zhang, D., Gu, Y., & Xing, D. (2021). Photoactivation of TGFβ/SMAD signaling pathway ameliorates adult hippocampal neurogenesis in Alzheimer’s disease model. Stem Cell Research & Therapy, 12(1), 345. https://doi.org/10.1186/s13287-021-02399-2
(15) Kandasamy, M., Lehner, B., Kraus, S., Sander, P. R., Marschallinger, J., Rivera, F. J., Trümbach, D., Ueberham, U., Reitsamer, H. A., Strauss, O., Bogdahn, U., Couillard-Despres, S., & Aigner, L. (2014). TGF-beta signalling in the adult neurogenic niche promotes stem cell quiescence as well as generation of new neurons. Journal of Cellular and Molecular Medicine, 18(7), 1444‑1459. https://doi.org/10.1111/jcmm.12298
(16) Johnston, S. T., Shtrahman, M., Parylak, S., Gonçalves, J. T., & Gage, F. H. (2016). Paradox of pattern separation and adult neurogenesis : A dual role for new neurons balancing memory resolution and robustness. Neurobiology of Learning and Memory, 129, 60‑68. https://doi.org/10.1016/j.nlm.2015.10.013
(17) Anacker, C., & Hen, R. (2017). Adult hippocampal neurogenesis and cognitive flexibility—Linking memory and mood. Nature Reviews Neuroscience, 18(6), 335‑346. https://doi.org/10.1038/nrn.2017.45
(18) Meng, C., He, Z., & Xing, D. (2013). Low-level laser therapy rescues dendrite atrophy via upregulating BDNF expression : Implications for Alzheimer’s disease. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 33(33), 13505‑13517. https://doi.org/10.1523/JNEUROSCI.0918-13.2013
